ডাইনোসর
ডাইনোসর বলতে জনপ্রিয় ধারণায় একটি অধুনা অবলুপ্ত, সাধারণত বৃহদাকার মেরুদণ্ডী প্রাণীগোষ্ঠীকে বোঝায়। এরা পৃথিবীর বাস্তুতন্ত্রের প্রাগৈতিহাসিক অধিবাসী এবং বৈজ্ঞানিকদের অনুমান এই প্রভাবশালী প্রাণীরা প্রায় ১৬ কোটি বছর ধরে পৃথিবীতে রাজত্ব করেছে। প্রথম ডাইনোসরের বিবর্তন হয়েছিল আনুমানিক ২৩ কোটি বছর পূর্বে। ক্রিটেশিয়াস যুগের শেষে প্রায় সাড়ে ৬ কোটি বছর পূর্বে একটি বিধ্বংসী প্রাকৃতিক বিপর্যয় ডাইনোসরদের প্রভাবকে পৃথিবী থেকে সম্পূর্ণ বিলুপ্ত করে দেয়। তাদের একটি শ্রেণীই কেবল বর্তমান যুগ পর্যন্ত টিকে থাকতে পেরেছে বলে ধারণা করা হয়: শ্রেণিবিন্যাসবিদরা ধারণা করেন আধুনিক পাখিরা থেরোপড ডাইনোসরদের সরাসরি বংশধর[১]; জীবাশ্ম দ্বারা প্রাপ্ত নিদর্শন থেকে জুরাসিক যুগে সংঘটিত এই বিবর্তনের প্রমাণ পাওয়া যায়[২]।
| ডাইনোসর সময়গত পরিসীমা: অন্ত্য ট্রায়াসিক–হলোসিন, ২৩.১৪–৬.৬ কোটি | |
|---|---|

| |
| কয়েকটি ডাইনোসর কঙ্কালের জীবাশ্ম। ঘড়ির কাঁটার গতিমুখ বরাবর একদম উপরে বাঁ দিক থেকে- মাইকোর্যাপ্টর গুই (একটি ডানাওয়ালা থেরোপড), | |
| বৈজ্ঞানিক শ্রেণীবিন্যাস | |
| জগৎ: | প্রাণী জগৎ |
| পর্ব: | কর্ডাটা |
| উপপর্ব: | মেরুদণ্ডী |
| শ্রেণী: | সরোপসিডা |
| উপশ্রেণী: | ডায়াপসিড |
| অধঃশ্রেণী: | আর্কোসরোমর্ফা |
| মহাবর্গ: | ডাইনোসরিয়া * ওয়েন, ১৮৪২ |
| বর্গ এবং উপবর্গ | |
শ্রেণিবিন্যাসগত, অঙ্গসংস্থানগত ও পরিবেশগত দিক থেকে ডাইনোসর কথাটিকে বিভিন্ন প্রকারের কতকগুলি প্রাণীর একটি সাধারণ নাম হিসেবে বর্ণনা করা যেতে পারে। জীবাশ্ম প্রমাণ থেকে পুরাজীববিদরা উড়তে অক্ষম ডাইনোসরদের ৫০০ এরও বেশি গণ[৩] ও ১০০০ এরও বেশি প্রজাতিকে শনাক্ত করেছেন[৪]। সব কয়টি মহাদেশেই ডাইনোসরদের জীবন্ত ও প্রস্তরীভূত নানা প্রজাতির দেখা পাওয়া যায়[৫], যাদের মধ্যে শাকাহারী ও মাংসাশী- উভয় প্রকার উদাহরণই রয়েছে। যদিও উৎপত্তিগতভাবে ডাইনোসরেরা দ্বিপদ, কিন্তু অবলুপ্ত অনেক চতুষ্পদ প্রজাতির সন্ধান পাওয়া গেছে, এবং কোনো কোনো প্রজাতি গমনের সময় প্রয়োজনমত দুই পা অথবা চার পা ব্যবহার করতে পারত। সমস্ত বিভাগের ডাইনোসরদের মধ্যেই শিং, হাড় ও চামড়ার পাত প্রভৃতি প্রদর্শনমূলক অঙ্গসংস্থানের নিদর্শন রয়েছে, এবং কোনো কোনো অবলুপ্ত প্রজাতির কঙ্কালে হাড়ের বর্ম ও কাঁটার মত গঠন লক্ষ্য করা যায়। বিভাগ নির্বিশেষে ডাইনোসরদের অন্যতম সাধারণ বৈশিষ্ট্য হল ডিম পাড়া ও বাসা বানানোর অভ্যাস। ওড়ার খাতিরে কিছু শারীরবৃত্তীয় বাধ্যবাধকতার জন্য আধুনিক পাখিরা আকারে ছোট হলেও প্রাগৈতিহাসিক ডাইনোসরদের অনেকেই ছিল বিশালদেহী। বৃহত্তম সরোপড ডাইনোসরেরা ৫৮ মিটার (১৯০ ফুট) পর্যন্ত দীর্ঘ এবং ৯.২৫ মিটার (৩০ ফুট ৪ ইঞ্চি) পর্যন্ত উঁচু হত[৬]। তবুও উড়তে অক্ষম ডাইনোসর মাত্রই বিশালাকার হবে- এই ধারণাটা ভুল। আবিষ্কৃত জীবাশ্মের বেশির ভাগই বড় মাপের ডাইনোসর- এ'কথা ঠিক। কিন্তু এর কারণ হল জীবাশ্মের আকার বড় হলে তা প্রকৃতির প্রতিকূলতা সহ্য করে প্রস্তরীভবন পর্যন্ত সহজে টিকে থাকতে পারে। আসলে অনেক ডাইনোসরই ছিল খুদে; যেমন, জিজিয়ানিকাস (Xixianykus) নামক ডাইনোসরটির দৈর্ঘ্য ছিল মাত্র ৫০ সেন্টিমিটার (প্রায় ২০ ইঞ্চি)।
যদিও 'ডাইনোসর' কথাটার আক্ষরিক অর্থ ভয়াবহ গিরগিটি, কিন্তু ডাইনোসরেরা প্রকৃতপক্ষে গিরগিটি নয়। বরং তারা সরীসৃপ শ্রেণীর অন্তর্গত একটা আলাদা গোষ্ঠীর প্রতিনিধি, যাদের শারীরবৃত্তীয় ক্রিয়াকলাপ অনেকাংশে বর্তমান সরীসৃপদের থেকে পৃথক; যেমন, তারা ছিল উষ্ণশোণিত এবং দ্বিপদ গমনে সক্ষম। বিংশ শতাব্দীর প্রথমার্ধে অর্থাৎ পাখিদের ডাইনোসর বলে চিহ্নিত করার আগে পর্যন্ত বৈজ্ঞানিকরা ডাইনোসরদের অলস এবং অনুষ্ণশোণিত বলে মনে করতেন। ১৯৭০ এর দশক এবং তৎপরবর্তী অধিকাংশ গবেষণা থেকে অবশ্য জানা গেছে যে সমস্ত ডাইনোসর ছিল উচ্চ বিপাক হার যুক্ত, অতিমাত্রায় সক্রিয় প্রাণী এবং তারা পরস্পরের সাথে যোগাযোগের জন্য বিভিন্নভাবে অভিযোজিত হয়েছিল।
ঊনবিংশ শতাব্দীর শুরুর দিকে ডাইনোসরের প্রথম জীবাশ্ম আবিষ্কৃত হয়। এরপর থেকে পর্বতগাত্র বা শিলায় আটকা পড়ে থাকা ডাইনোসরের কঙ্কাল পৃথিবীর বিভিন্ন জাদুঘরে আকর্ষণের কেন্দ্রবিন্দুতে পরিণত হয়। ডাইনোসরেরা বর্তমান বিশ্ব সংস্কৃতির একটি অবিচ্ছেদ্য অঙ্গে পরিণত হয়েছে। প্রধানত কোনো কোনো অবলুপ্ত ডাইনোসর প্রজাতির বিশাল আয়তন এবং তাদের সম্ভাব্য হিংস্র স্বভাবের দরুন তারা শিশু ও বয়স্ক সবার কাছেই বিশেষ আগ্রহের বিষয়ে পরিণত হয়েছে। সর্বাধিক বিক্রিত বই এবং জুরাসিক পার্ক ইত্যাদি প্রচুর কাটতি পাওয়া চলচ্চিত্রে ডাইনোসর প্রসঙ্গ এসেছে এবং এ সংক্রান্ত নতুন যে কোনো আবিষ্কার গণমাধ্যমে বিশেষভাবে সম্প্রচার করা হচ্ছে।
ব্যুৎপত্তি
সম্পাদনা১৮৪২ খ্রীষ্টাব্দে পুরাজীববিদ স্যার রিচার্ড ওয়েন, ডাইনোসরিয়া ট্যাক্সনটির আনুষ্ঠানিক নামকরণ করেন। তৎকালীন ইংল্যান্ড এবং অবশিষ্ট পৃথিবীর নানা অঞ্চল থেকে সরিয়ান (Saurian) গোষ্ঠীর সরীসৃপদের যে "বিশিষ্ট শাখা অথবা উপবিভাগের" সদস্যদের অবশেষ খুঁজে পাওয়া যাচ্ছিল, তাদের নির্দিষ্ট করতেই এই শব্দটির ব্যবহার শুরু হয়[৭]। গ্রিক ভাষার δεινός (দেইনস, অর্থাৎ "ভীষণ" বা "ভয়াবহ") এবং σαῦρος (সাউরোস, অর্থাৎ "গোধা" বা "গিরগিটি")- এই দু'টি শব্দ জুড়ে ডাইনোসর কথাটি তৈরী হয়েছে[৭][৮]। যদিও নামটা ডাইনোসরদের দাঁত, নখ ও অন্যান্য ভয়াবহ বৈশিষ্ট্যের কথা মনে পড়িয়ে দেয়, কিন্তু ওয়েন এই নামকরণ করেছিলেন প্রধানত প্রাণীগুলোর আয়তনের কথা মাথায় রেখে[৯]।
ব্যবহৃত পরিভাষা
সম্পাদনা| বাংলা | ইংরেজি |
|---|---|
| পক্ষিকুল | Aves |
| মেরুদণ্ডী | Vertebrata |
| জাতিজনি শ্রেণিবিন্যাস | Phylogenetic taxonomy |
| স্থল গমন | Terrestrial locomotion |
| বাস্তুতন্ত্রগত ধাপ | Ecological niche |
সংজ্ঞা
সম্পাদনা
জাতিজনি শ্রেণিবিন্যাসের অধীনে সাধারণত ট্রাইসেরাটপস, নিঅর্নিথিস (আধুনিক পাখি), এদের সাম্প্রতিকতম সাধারণ পূর্বপুরুষ(সা.সা.পূ./Most Recent Common Ancestor/MRCA) ও তাদের সমস্ত বংশধরকে ডাইনোসর বলা হয়[১০]। অন্য একটি মত অনুযায়ী মেগালোসরাস এবং ইগুয়ানোডন এর সা.সা.পূ. থেকে ডাইনোসরদের চিহ্নিত করার প্রক্রিয়া শুরু করা উচিত, কারণ রিচার্ড ওয়েন যখন ডাইনোসরিয়া-র শনাক্তকরণ করেছিলেন তখন এই দু'টি ছিল তার দ্বারা নির্দিষ্ট তিনটি গণের অন্যতম[১১]। দু'টি পদ্ধতিই একই জীবসমষ্টিকে ডাইনোসর হিসেবে চিহ্নিত করে: ডাইনোসরিয়া= অর্নিথিস্কিয়া+সরিস্কিয়া; যাদের অন্তর্গত হল থেরোপড (প্রধানত দ্বিপদ মাংসাশী এবং পাখি), অ্যাঙ্কিলোসর (বর্মযুক্ত শাকাহারী চতুষ্পদ), স্টেগোসর (পাতযুক্ত শাকাহারী চতুষ্পদ), সেরাটপ্সিয়া (শিং ও শিরস্ত্রাণযুক্ত শাকাহারী চতুষ্পদ), অর্নিথোপড (হাঁসের মত চঞ্চুবিশিষ্ট শাকাহারী দ্বিপদ বা চতুষ্পদ) এবং সরোপডোমর্ফ (প্রধানত লম্বা গলা ও লেজযুক্ত বৃহৎ শাকাহারী চতুষ্পদ) রা[১২]।

বর্তমানে পাখিদেরকে থেরোপড ডাইনোসরদের একমাত্র জীবিত বংশধররূপে গণ্য করা হয়। গতানুগতিক শ্রেণিবিন্যাসবিদ্যায় পাখিদের ডাইনোসর থেকে বিবর্তিত একটা আলাদা শ্রেণী বলে মনে করা হত। অবশ্য ডাইনোসর নিয়ে আগ্রহী সাম্প্রতিক জীববৈজ্ঞানিকদের অধিকাংশই গতানুগতিক পদ্ধতির পরিবর্তে জাতিজনি নামকরণের পক্ষ সমর্থন করেন। এই পদ্ধতি অনুযায়ী কোনো প্রাণী বা উদ্ভিদ বিভাগের সমস্ত উত্তরপুরুষ ঐ বিভাগের সদস্য হিসেবে গণ্য হয়। পাখিরা তাই ডাইনোসর,আর সেই সূত্রেই ডাইনোসরেরা বিলুপ্ত নয়, জীবিত। পাখিদের ম্যানির্যাপ্টোরা অধঃবিভাগের অন্তর্গত মনে করা হয়; ম্যানির্যাপ্টোরা বর্গটি সিলুরোসরাসদের অন্তর্গত; সিলুরোসরাসরা এক প্রকার থেরোপড, থেরোপডরা এক প্রকার সরিস্কিয়ান এবং সরিস্কিয়ানরা এক প্রকার ডাইনোসর[১৩]। কিন্তু সাধারণের বক্তব্য ধর্তব্যের মধ্যে আনলে ডাইনোসরের মধ্যে পাখিদেরকে বাদ দিতে হয়। স্পষ্টতার খাতিরে এই নিবন্ধে, "ডাইনোসর" শব্দটি বলতে "উড়তে অক্ষম ডাইনোসর"-দের বোঝানো হবে এবং "পাখি" শব্দটি "উড়তে সক্ষম ডাইনোসর"-এর প্রতিশব্দ হিসেবে ব্যবহৃত হবে। উড়তে সক্ষম ডাইনোসর বলতে আর্কিওপ্টেরিক্স পূর্বপুরুষ থেকে বিবর্তিত এবং আধুনিক পাখিদের সবাইকেই বোঝানো হবে। গুরুত্ব দিয়ে কোন বিষয় উল্লেখ করতে হলে "উড়তে অক্ষম ডাইনোসর" শব্দটিই ব্যবহৃত হবে।
সাধারণ বর্ণনা
সম্পাদনাউপরে প্রদত্ত একটি সংজ্ঞা ব্যবহার করে বলা যায়, দেহের সম্পূর্ণ ভার বহনক্ষম উপাঙ্গ যুক্ত আর্কোসরেরা হল ডাইনোসর[১৪]। ডাইনোসর নামটি জনপ্রিয় ধারণায় অন্যান্য কিছু প্রাগৈতিহাসিক সরীসৃপকে বোঝাতেও ব্যবহৃত হয়। এর মধ্যে রয়েছে ডিমেট্রোডন নামক পেলিকোসর, পাখাবিশিষ্ট টেরোসর, জলচর প্লেসিওসর এবং মোসাসর। কিন্তু প্রকৃতপক্ষে এগুলোর কোনটিই ডাইনোসর ছিল না এবং এদের কারোরই প্রকৃত ডাইনোসরদের মত সম্পূর্ণ দেহভার বহনকারী উপাঙ্গ ছিল না[১৫]। ডাইনোসরেরা ছিল মেসোজোয়িক মহাযুগের, বিশেষত জুরাসিক ও ক্রিটেশিয়াস কালপর্বের প্রধান স্থলচর মেরুদণ্ডী। অন্যান্য ধরনের প্রাণীদের প্রভাব আয়তন ও বাস্তুতন্ত্রগত ধাপের নিরিখে ছিল খুব সীমিত। যেমন, সমসাময়িক স্তন্যপায়ীরা প্রায় কখনোই আয়তনে আধুনিক বিড়ালের থেকে বড় হত না; তাদের বেশির ভাগই ছিল ইঁদুর জাতীয় ক্ষুদ্র পতঙ্গভুক জীব[১৬]।

ডাইনোসরেরা বরাবরই বিচিত্র এবং বহু-বিভাজিত একটি প্রাণীগোষ্ঠী হিসেবে বিবেচিত হয়েছে। ২০০৬ খ্রীষ্টাব্দের একটি সমীক্ষা অনুযায়ী উড়তে অক্ষম ডাইনোসরদের ৫০০ এরও বেশি গণ এখনও অবধি নিশ্চিতভাবে শনাক্ত করা গেছে, এবং আন্দাজ করা হয় যে সারা পৃথিবীতে সঞ্চিত মোট ডাইনোসর জীবাশ্মের মধ্যে প্রায় ১৮৫০ টির কাছাকাছি সংখ্যক আলাদা আলাদা গণ রয়েছে, যার প্রায় ৭৫ শতাংশ আজও আবিষ্কার করা যায়নি[১৭]। এতদপেক্ষা কিঞ্চিৎ পুরোনো একটি গবেষণায় ডাইনোসরদের গণের সংখ্যা ৩৪০০ নির্ধারণ করা হয়েছিল, যার মধ্যে অনেকগুলিই সম্ভবত জীবাশ্মে পরিণত হতে পারে নি[১৮]। ২০০৮ খ্রীষ্টাব্দের ১৭ই সেপ্টেম্বর পর্যন্ত ১০৪৭ টি ভিন্ন ভিন্ন ডাইনোসর প্রজাতি আবিষ্কৃত হয়েছে[১৯]। এদের মধ্যে কেউ কেউ শাকাহারী এবং কেউ কেউ মাংসাশী। ডাইনোসরদের বিভিন্ন প্রকার খাদ্যাভ্যাসের মধ্যে গাছের বীজ,মাছ, পতঙ্গ ইত্যাদি নানা উপাদানের সমাবেশ ছিল; এমনকি সর্বভুক ডাইনোসরের নিদর্শনও পাওয়া গেছে। যদিও ডাইনোসরেরা (এবং সেইসূত্রে পাখিরা) উৎপত্তিগতভাবে দ্বিপদ, কোনো কোনো প্রাগৈতিহাসিক প্রজাতি চতুষ্পদও ছিল। এছাড়া অ্যামোসরাস এবং ইগুয়ানোডনের মত কিছু প্রজাতি ইচ্ছামত গমনের সময় চার পা বা দুই পা ব্যবহার করতে পারত। জীবাশ্ম প্রমাণ থেকে জানা যায়, আদি জুরাসিক যুগেই ডাইনোসরেরা সারা পৃথিবীতে ছড়িয়ে পড়েছিল[২০]। আধুনিক পাখিদের আবাসস্থল বহুবৈচিত্র্যপূর্ণ; তারা স্থল ও সামুদ্রিক- উভয় পরিবেশেই স্বচ্ছন্দ থাকতে পারে। এছাড়া মাইকোর্যাপ্টর এর মত কিছু কিছু উড়তে অক্ষম ডাইনোসরের বাতাসে ভর করে সাময়িকভাবে ভাসমান থাকার ক্ষমতা এবং স্পিনোসরিড দের আংশিক জলচর স্বভাবের পক্ষেও প্রমাণ আছে[২১]।
গুরুত্বপূর্ণ শারীরস্থানিক বৈশিষ্ট্য
সম্পাদনাযদিও সাম্প্রতিক গবেষণাসমূহের পর ডাইনোসরদের শারীরস্থানিক বৈশিষ্ট্যের কোনো সর্বজনসম্মত তালিকা প্রকাশ করা দুরূহ হয়ে উঠেছে, তবুও আজ পর্যন্ত আবিষ্কৃত অধিকাংশ ডাইনোসর হয় আদিম আর্কোসরীয় কঙ্কালের কিছু গঠনগত বৈশিষ্ট্য দেখায়, নয়তো ঐ সমস্ত বৈশিষ্ট্যযুক্ত কোনো পূর্বপুরুষ থেকেই উদ্ভূত হয়েছে বলে বোঝা যায়। বিবর্তনের ধারায় অপেক্ষাকৃত পরের দিকে আসা কিছু প্রজাতির দেহে এই বৈশিষ্ট্যগুলি বেশি মাত্রায় পরিবর্তিত হয়ে গেছে। এই বৈশিষ্ট্যগুলি ডাইনোসরিয়ার সাধারণ চরিত্রগত বলে মনে করা হয়; প্রাচীনতম ডাইনোসরেরা এ'গুলি তাদের বংশধরদের দেহে সঞ্চারিত করে গেছে। এই ধরনের বৈশিষ্ট্য-সমষ্টিগুলি কোনো নির্দিষ্ট শ্রেণিবিন্যাসগত বিভাগের সাম্প্রতিকতম সাধারণ পূর্বপুরুষের দেহে উৎপন্ন হয় এবং এগুলিকে ঐ নির্দিষ্ট বিভাগের সাইন্যাপোমর্ফি (synapomorphies) বলে[২২]।
আর্কোসরদের আন্তঃসম্পর্ক সম্বন্ধে এস নেসবিট[২৩]-এর পুঙ্খানুপুঙ্খ গবেষণা নিম্নলিখিত বারোটি সাইন্যাপোমর্ফি নির্ভুলভাবে চিহ্নিত করেছে। কোনো কোনোটা আগে থেকেই জানা ছিল:



- মাথার খুলির ছাদের পিছন দিকের দু'টি প্রধান গহ্বরের (সুপ্রাটেম্পোরাল ফেনেস্ত্রা/Supra-temporal Fenestra) সামনে একটি করে অবতল খাঁজ বা ফসা থাকে।
- ঘাড়ের প্রথম দু'টি কশেরুকা অ্যাটলাস এবং অ্যাক্সিস-এর সামনের দিকে এপিপোফাইসিস নামক উপবৃদ্ধি দেখা যায়।
- ডেল্টো-পেক্টোরাল পেশিসমূহের সংযোগস্থল অগ্রপদের প্রথম হাড় প্রগণ্ডাস্থির ৩০% বা তার বেশি দৈর্ঘ্যে অবস্থিত।
- অগ্রপদের রেডিয়াস হাড়ের দৈর্ঘ্য হিউমেরাস বা প্রগণ্ডাস্থির ৮০% এর কম।
- উরুর হাড়ের পিছন দিকে কডোফিমোরালিস পেশির সংযোগস্থলে অবস্থিত উপবৃদ্ধিটি (চতুর্থ ট্রোক্যান্টার) অতি স্পষ্ট এবং কানা-উঁচু চামচের আকৃতিবিশিষ্ট (ছবি দেখুন)।
- চতুর্থ ট্রোক্যান্টার আকারে অসমান; এর এক দিকের কানা অন্য দিক অপেক্ষা বেশি উঁচু থাকে।
- গোড়ালির অস্থিসমূহের সাথে পায়ের অনুজঙ্ঘাস্থি হাড়ের প্রথম সংযোগস্থলটি ঐ হাড়গুলির ৩০% এর কম স্থান অধিকার করে থাকে।
- খুলির পিছন দিকে অবস্থিত বহিঃঅক্সিপিটাল হাড় দু'টি মস্তিষ্ক-গহ্বরের মেঝের মাঝ বরাবর পরস্পরের সাথে মিলিত হয় না।
- শ্রোণীচক্রের ইস্কিয়াম হাড়টির ভিতর দিকে অবস্থিত একটি বৃহৎ অবতল খাঁজ ইলিয়াম ও পিউবিস হাড়ের থেকে এটিকে পৃথক রাখে।
- জঙ্ঘাস্থি বা টিবিয়ার সামনে উপর দিকে পেশির সংযোগের জন্য যে উপবৃদ্ধি (নেমিয়াল ক্রেস্ট) থাকে, সেটি বাইরের দিকে বাঁকানো হয়।
- জঙ্ঘাস্থির নিচের অংশে পিছন দিকে একটি উল্লম্ব পাতের আকারের উপবৃদ্ধি থাকে।
- অনুজঙ্ঘাস্থি বা ফিবুলার যে তলটি ক্যালকেনিয়াস হাড় অর্থাৎ গোড়ালি অঞ্চলের প্রথম হাড়ের সাথে যুক্ত থাকে, সেটি অবতল।
নেসবিট আরও কতকগুলি সম্ভাব্য সাইন্যাপোমর্ফি খুঁজে পেয়েছিলেন, এবং পূর্বে উল্লিখিত কতকগুলি সাইন্যাপোমর্ফি তার তালিকা থেকে বাদ দিয়েছিলেন। তার তালিকার কোনো কোনো বৈশিষ্ট্য সিলেসরিডদের মধ্যেও দেখা যায়, যাদেরকে তিনি ডাইনোসরদের জ্ঞাতি হিসেবে পুনঃচিহ্নিত করেন। ডাইনোসর ও সিলেসরিডদের সাধারণ সাইন্যাপোমর্ফির মধ্যে ছিল সম্মুখবর্তী বৃহৎ ট্রোক্যান্টার, দ্বিতীয় ও চতুর্থ মেটাটার্সাল অস্থির অসমান দৈর্ঘ্য, ইস্কিয়াম এবং পিউবিসের সীমিত সংযোগস্থল, টিবিয়ায় নেমিয়াল ক্রেস্টের উপস্থিতি, ট্যালাস হাড়ে একটি ঊর্দ্ধমুখী উপবৃদ্ধি এবং আরও অনেক[২৪]।

ডাইনোসরেরা অধিকাংশ আধুনিক স্তন্যপায়ীর মতো খাড়া হয়ে দাঁড়াতে পারত, কারণ তাদের শ্রোণীচক্র গঠনগতভাবে অন্যান্য স্থলচর সরীসৃপদের (যারা হামাগুড়ি দিয়ে চলে) থেকে আলাদা ছিল[২৫]। এর কারণ হিসেবে শ্রোণীচক্রের একটি আনুভূমিক উপবৃদ্ধি এবং ঊর্বস্থি বা ফিমারের অভ্যন্তরমুখী মস্তকের বিবর্তনকে দায়ী করা যায়[২৬]। সোজা হয়ে দাঁড়ানোর ক্ষমতা প্রথম ডাইনোসরদের শ্বাসকার্যের ক্ষেত্রে বাড়তি সুবিধে দিয়েছিল, যার ফলে তাদের পরিশ্রম করার ক্ষমতা সাধারণ 'ছেতরে থাকা' পা-ওয়ালা সরীসৃপদের চেয়ে অনেকটা বৃদ্ধি পায়[২৭]। ছেতরে থাকা পায়ের উপর যে অতিরিক্ত চাপ পড়ে সেটা এড়াতে পারার ফলে সোজা হয়ে থাকা পা সম্ভবত দেহের বিশাল আয়তনেরও অনুকূল হয়েছিল[২৮]। রাউইসুচিয়া সমেত কোনো কোনো অ-ডাইনোসর আর্কোসরও সোজা হয়ে দাঁড়ানোর ক্ষমতা আয়ত্ত করেছিল, কিন্তু তাদের ক্ষেত্রে এটা সম্ভব হয়েছিল শ্রোণীচক্রের ঋজু-স্তম্ভাকার গঠনের ফলে, যেখানে ঊর্বস্থির গোলাকার মস্তক শ্রোণীর গর্তের মধ্যে সংযুক্ত থাকার পরিবর্তে শ্রোণীচক্রের উপরের হাড়টি প্রলম্বিত হয়ে ঊর্বস্থি দু'টিকে খাড়া রাখত[২৮]।
বিবর্তনীয় ইতিহাস
সম্পাদনামূল নিবন্ধː ডাইনোসরের বিবর্তন
উৎপত্তি এবং প্রাথমিক বিবর্তন
সম্পাদনা

পার্মিয়ান-ট্রায়াসিক অবলুপ্তি ঘটনায় পৃথিবীর জীববৈচিত্র্যের প্রায় ৯৫% ধ্বংস হয়ে যাওয়ার কমবেশি ২ কোটি বছর পর মধ্য থেকে অন্ত্য ট্রায়াসিকে ডাইনোসরেরা তাদের আর্কোসর পূর্বপুরুষদের থেকে পৃথক হয়ে যায়[২৯][৩০]। তেজস্ক্রিয়মিতিক তারিখ গণনার মাধ্যমে ২৩ কোটি ১৪ লক্ষ বছর আগেকার পাথরের স্তরে প্রাথমিক ডাইনোসর গণ ইওর্যাপ্টরের জীবাশ্ম আবিষ্কৃত হয়েছে[৩১]। পুরাজীববিদরা ধারণা করেন ইওর্যাপ্টর সমস্ত ডাইনোসরের সাধারণ পূর্বপুরুষের নিকট জ্ঞাতি[৩২]। এই ধারণা সত্যি হলে বলা যেতে পারে যে প্রথম ডাইনোসরেরা ছিল আকারে অপেক্ষাকৃত ছোট দ্বিপদ মাংসাশী প্রাণী[৩৩]। আর্জেন্টিনার মধ্য ট্রায়াসিক পাথরের স্তর থেকে মারাসুকাস এবং ল্যাগারপেটন নামক আদিম, ডাইনোসর-সদৃশ অর্নিথোডিরানদের আবিষ্কার এই ধারণার ভিত্তি দৃঢ় করে, কারণ এরাও ছিল ছোট, দ্বিপদ এবং মাংসভুক। নিয়াসাসরাসের জীবাশ্ম ইঙ্গিত করে ডাইনোসরেরা হয়তো ২৪ কোটি ৩০ লক্ষ বছর আগেই বিবর্তিত হয়েছিল, যদিও জীবাশ্মটির খুব অল্প অংশ উদ্ধার করা গেছে বলে এরা সত্যিই ডাইনোসর ছিল নাকি ডাইনোসরদের কোনো নিকটাত্মীয় সরীসৃপ ছিল তা নিয়ে নিশ্চিত হওয়া যায় না[৩৪]।
ডাইনোসরদের আবির্ভাবের সময় উল্লেখযোগ্য স্থলচর জীবগোষ্ঠীর মধ্যে ছিল নানা জাতের আর্কোসরোমর্ফ এবং থেরাপসিড সরীসৃপ, যথা এইটোসর, কাইনোডন্ট, অর্নিথোসুকিডি, রাউইসুকিয়া এবং রিঙ্কোসর। পর পর দু'টো অবলুপ্তি ঘটনায় এই গোষ্ঠীগুলোর বেশিরভাগই লুপ্ত হয়ে যায়। প্রথমত ২১ কোটি ৫০ লক্ষ বছর আগে প্রোটোরোসর সমেত বহু প্রজাতির আর্কোসরোমর্ফের বিলুপ্তি ঘটে। এর পর ট্রায়াসিক-জুরাসিক অবলুপ্তি ঘটনায় (২০ কোটি বছর আগে) এইটোসর, অর্নিথোসুকিডি, ফাইটোসর প্রভৃতি আর্কোসর এবং রাউইসুকিয়ানরা অবলুপ্ত হয়। রিঙ্কোসর এবং ডিকাইনোডন্টরা অন্ত্য ট্রায়াসিকের শেষভাগ অবধি টিকে ছিল[৩৫][৩৬] এবং এদের অবলুপ্তির সঠিক সময় সম্বন্ধে এখনও নিশ্চিত হওয়া যায়নি। এই সমস্ত অবলুপ্তি ঘটনার ফলে ডাঙার প্রাণী হিসেবে অবশিষ্ট রইল ক্রোকোডাইলোমর্ফ, ডাইনোসর, স্তন্যপায়ী, টেরোসর এবং কচ্ছপেরা[১০]। ডাইনোসরেদের প্রথম গোষ্ঠীগুলো ট্রায়াসিকের কার্নিয়ান ও নরিয়ান পর্যায়ে নানা শাখায় ভাগ হয়ে গিয়ে ক্রমশ অবলুপ্ত প্রজাতিগুলোর জায়গা দখল করে[১২]।
বিবর্তন ও পুরাজীবভূগোল
সম্পাদনাট্রায়াসিক যুগের পর থেকে ডাইনোসরদের বিবর্তন স্বাভাবিক উদ্ভিদ ও মহাদেশসমূহের প্রকৃতি ও অবস্থান পরিবর্তনের সাথে তাল মিলিয়ে হয়েছিল। অন্ত্য ট্রায়াসিক ও আদি জুরাসিকে সমস্ত মহাদেশ একত্রে প্যাঞ্জিয়া নামক একটা অখণ্ড অতিমহাদেশ গঠন করে ছিল, আর পৃথিবীব্যাপী ডাইনোসরেরা প্রধানত সিলোফাইসিড জাতীয় মাংসাশী এবং আদি সরোপডোমর্ফ জাতীয় শাকাহারী এই দুই ভাগে বিভক্ত ছিল[৩৭]। ডাইনোসরদের সম্ভাব্য খাদ্য কনিফারপ্রধান ব্যক্তবীজী উদ্ভিদগোষ্ঠী অন্ত্য ট্রায়াসিকে নানা প্রজাতিতে ভাগ হয়ে যায়। প্রাথমিক সরোপডোমর্ফদের চোয়াল তথা মুখবিবরের গঠন তাদের খাবার থেঁতলে হজমের উপযোগী করে তোলার উপযুক্ত ছিল না। তাই ধারণা করা হয় তারা খাদ্যনালীর পরবর্তী কোনো অংশে এই কাজগুলো সম্পন্ন করার ব্যবস্থা করেছিল[৩৮]। ডাইনোসরদের প্রজাতিগত বৈচিত্র্যহীনতা মধ্য থেকে অন্ত্য জুরাসিক পর্যন্ত বজায় ছিল এবং এই গোটা সময় জুড়ে ভূখণ্ডের যে কোনো অঞ্চলে মাংসাশী বলতে ছিল সেরাটোসরিয়া, স্পিনোসরিড ও কার্নোসররা আর শাকাহারীদের মধ্যে ছিল স্টেগোসর জাতীয় অর্নিথিস্কিয়ান ও বৃহদায়তন সরোপডরা। উত্তর আমেরিকার মরিসন প্রস্তরক্ষেত্র এবং তাঞ্জানিয়ার তেন্দাগুরু থেকে এই জীববণ্টনের প্রমাণ মেলে। চীনের ডাইনোসররা একটু আলাদা ছিল; সিনর্যাপ্টর প্রভৃতি থেরোপড এবং মামেঞ্চিয়াসরাস ইত্যাদি বিশেষ প্রকার সরোপডরা এখানকার বাসিন্দা ছিল[৩৭]। অ্যাঙ্কিলোসর এবং অর্নিথোপডরা সংখ্যায় বাড়ছিল, কিন্তু প্রোসরোপডরা বিলুপ্ত হয়ে গিয়েছিল। পাইন তথা কনিফার এবং ফার্ন ছিল প্রধান উদ্ভিদ। আগেকার প্রোসরোপডদের মত সরোপডরাও গিলে ফেলা ছাড়া মুখের মধ্যে খাবারের কোনো রকম পাচন-সহায়ক কাজ করত না, কিন্তু অর্নিথিস্কিয়ানরা এই ব্যাপারে অনেক দূর এগিয়ে গিয়েছিল; খাবার অনেকক্ষণ ধরে মুখের মধ্যে নিয়ে চিবানোর সুবিধার জন্য তাদের মধ্যে কারো কারো মুখে গাল সদৃশ অঙ্গেরও আবির্ভাব ঘটেছিল, আর চিবানোর কাজ আরও নিখুঁত করার জন্য দেখা দিয়েছিল নমনীয় চোয়াল[৩৮]। জুরাসিক যুগে বিবর্তনের আর একটা উল্লেখযোগ্য ঘটনা হল ম্যানির্যাপ্টোরা জাতীয় সিলুরোসরিডদের থেকে প্রথম পাখিদের আবির্ভাব[৩৯]।
আদি ক্রিটেশিয়াস নাগাদ প্যাঞ্জিয়া অতিমহাদেশ ভেঙে গিয়ে তার বিভিন্ন টুকরোগুলো একে অপরের থেকে দূরে সরে যাওয়ায় বাসস্থানের স্বতন্ত্র বৈশিষ্ট্য অনুযায়ী ডাইনোসরেরা নানা শাখায় ভাগ হয়ে যায়। এই সময়ে অ্যাঙ্কিলোসর, ইগুয়ানোডন্টিয়া এবং ব্র্যাকিওসরিডরা ইউরোপ, উত্তর আমেরিকা ও উত্তর আফ্রিকা জুড়ে বিচরণ করত। পরবর্তীকালে এদের জায়গা নেয় স্পিনোসরিড ও কার্কারোডন্টোসরিড জাতীয় থেরোপড এবং রেবাচিসরিড ও টাইটানোসর জাতীয় সরোপড; শেষোক্ত সরোপডদের জীবাশ্ম দক্ষিণ আমেরিকাতেও পাওয়া গেছে। এশিয়ায় ড্রোমিওসরিড, ট্রুডন্টিড ও ওভির্যাপ্টরোসরিয়ারা ছিল প্রধান থেরোপড এবং শাকাহারীদের মধ্যে উল্লেখযোগ্য ছিল অ্যাঙ্কিলোসর ও সিটাকোসরাস প্রভৃতি প্রাথমিক সেরাটোপ্সিয়ান। অন্যদিকে অস্ট্রেলিয়ায় প্রাধান্য ছিল প্রাথমিক অ্যাঙ্কিলোসর, হিপসিলোফোডন্টিড এবং ইগুয়ানোডন্টিয়াদের[৩৭]। ধারণা করা হয় আদি ক্রিটেশিয়াসের শেষভাগে স্টেগোসরিয়া জাতীয় ডাইনোসরেরা বিলুপ্ত হয়ে গিয়েছিল। আদি ক্রিটেশিয়াসের আরও একটা প্রধান ঘটনা হল সপুষ্পক উদ্ভিদের বিবর্তন, যারা অন্ত্য ক্রিটেশিয়াসে খুব তাড়াতাড়ি নানা প্রজাতিতে ভাগ হয়ে যায়। এর সাথে তাল মিলিয়ে শাকাহারী ডাইনোসরেরা গাছপালা চিবিয়ে হজম করার আলাদা আলাদা কায়দা রপ্ত করছিল। সেরাটোপ্সিয়ানরা একটার উপর একটা দাঁত একসাথে জুড়ে গিয়ে তৈরি যৌগিক দাঁতের মাধ্যমে খাবার টুকরো করতে পারত আর ইগুয়ানোডন্টরা তাদের অন্য রকম যৌগিক দাঁত ব্যবহার করত খাবার থেঁতো করার কাজে; এই শেষোক্ত পদ্ধতির চূড়ান্ত উন্নতি দেখা যায় হ্যাড্রোসরিডদের জীবাশ্মে[৩৮]। নাইজারসরাস এর মত কোনো কোনো সরোপডেরও যৌগিক দাঁত ছিল[৪০]।
সামগ্রিকভাবে বলা যায় অন্ত্য ক্রিটেশিয়াসে তিনটি স্বতন্ত্র পরিবেশে ডাইনোসরদের বাস ছিল। উত্তর আমেরিকা ও এশিয়ায় প্রধান থেরোপড বলতে ছিল টির্যানোসরিড এবং অন্যান্য ম্যানির্যাপ্টোরান প্রজাতি আর শাকাহারী বলতে ছিল প্রধানত নানা প্রকার অর্নিথিস্কিয়ান যথা হ্যাড্রোসরিড, সেরাটোপ্সিয়ান, অ্যাঙ্কিলোসর এবং প্যাকেসেফালোসরিয়ানরা। ভেঙে যাওয়া গন্ডোয়ানা থেকে সৃষ্ট দক্ষিণের মহাদেশগুলোয় অ্যাবেলিসরিডরা ছিল প্রধান থেরোপড আর টাইটানোসর জাতীয় সরোপডরা ছিল প্রধান শাকাহারী। তৃতীয়ত ইউরোপে ড্রোমিওসরিডরা ছিল প্রধান থেরোপড, আর র্যাবোডন্টিড জাতীয় ইগুয়ানোডন, নোডোসরিড জাতীয় অ্যাঙ্কিলোসর এবং টাইটানোসরীয় সরোপডদের প্রাধান্য ছিল[৩৭]। সপুষ্পক উদ্ভিদদের অনেক প্রজাতির আবির্ভাব হয়েছিল[৩৮]; এদেরই অন্যতম শাখা হিসেবে ঘাসের বিবর্তন হয় ক্রিটেশিয়াসের একেবারে অন্তিম লগ্নে[৪১]। উত্তর আমেরিকা ও এশিয়া জুড়ে সেরাটোপ্সিয়ান ও হ্যাড্রোসরিডদের অজস্র প্রজাতির বিবর্তন হচ্ছিল। এমনকি থেরোপডেরা পর্যন্ত থেরিজিনোসর ও অর্নিথোমিমোসরিয়া প্রভৃতি শাকাহারী বা সর্বভুক প্রজাতির জন্ম দেয়[৩৮]।
৬.৬ কোটি বছর আগে সংঘটিত ক্রিটেশিয়াস-প্যালিওজিন বিলুপ্তি ঘটনায় নিঅর্নিথিন পাখি ছাড়া সমস্ত ডাইনোসর বিলুপ্ত হয়ে যায়। সরীসৃপদের মধ্যে অন্য কোনো কোনো ডায়াপসিড বিভাগ যেমন কুমির, সেবেকোসুকিয়া, কচ্ছপ, গিরগিটি, সাপ, স্ফেনোডন্ট এবং করিস্টোডেরারা ঐ ঘটনার পরেও টিকে থাকতে সমর্থ হয়েছিল[৪২]।
আধুনিক দৌড়বাজ পাখি, হাঁস-মুরগি এবং নানা প্রকার জলের পাখিরা প্যালিওজিন যুগে দ্রুত বিভাজিত হয়ে পরিবেশে ডাইনোসরদের ছেড়ে যাওয়া স্থান দখল করে নেয়। এদের কয়েকটি প্রথম শাখা হল গেছো এনান্টিঅর্নিথিন, জলের হেস্পারর্নিথিন, এবং বিরাট আয়তনের গ্যাস্টর্নিস, মিহিরুং ও "ভয়াল পাখি" প্রভৃতি উড়তে অক্ষম স্থলচর পাখি। অবশ্য স্তন্যপায়ীরাও খুব তাড়াতাড়ি বিভাজিত হচ্ছিল আর তারাই ক্রমশ শুকনো ডাঙায় পাখিদের চেয়ে বেশি প্রভাবশালী হয়ে ওঠে[৪৩]।
ডাইনোসরের শ্রেণিবিন্যাস
সম্পাদনাআধুনিক কুমিরদের মত ডাইনোসরেরাও আর্কোসর জাতীয় সরীসৃপদের প্রকারবিশেষ। অন্যান্য আর্কোসরেদের থেকে ডাইনোসরদের গমনের ধরন ছিল আলাদা। কুমির ও টিকটিকি, গিরগিটিদের পা দেহের দু'পাশে ছেতরে থাকলেও ডাইনোসরদের পশ্চাৎপদ দেহের নিচে সোজা হয়ে থাকত[৪৪]।
সামগ্রিকভাবে জীবশাখা হিসেবে ডাইনোসরেরা প্রধান দু'টো শাখায় বিভক্ত, সরিস্কিয়া এবং অর্নিথিস্কিয়া। সরিস্কিয়াদের অন্তর্গত হল সেই সমস্ত প্রজাতি যারা অর্নিথিস্কিয়াদের চেয়ে পাখিদের নিকটতর জ্ঞাতি, আর অর্নিথিস্কিয়া হল তারা যারা সরিস্কিয়াদের চেয়ে ট্রাইসেরাটপ্স দের সাথে নিকটতর সম্পর্কযুক্ত। শারীরস্থানিকভাবে এই দু'টো বিভাগের সদস্যদের তাদের শ্রোণীচক্রের গঠনের তারতম্য দেখে চিহ্নিত করা যায়।
সরিস্কিয়া—যাদের নামের আক্ষরিক অর্থ "গিরগিটির মত কোমর" (গ্রিক σαῦρος/"সাউরোস" বা সরীসৃপ এবং ἰσχίον/"ইস্কিয়ন" বা পশ্চাদ্দেশ)—দের দেহে তাদের পূর্বপুরুষদের শ্রোণীচক্রের গঠন অপরিবর্তিত ছিল; এদের পিউবিস হাড় সামনের দিকে বাঁকানো ছিল[৪৫]। সরিস্কিয়াদের বিভিন্ন প্রজাতিতে এই বুনিয়াদী গঠন অপরিবর্তিত ছিল, কেবল পিউবিস হাড়ের আগাটা প্রজাতিভেদে বিভিন্ন কোণে পিছনের দিকে বাঁক নিতে দেখা যায় (হেরেরাসরাস[৪৬], থেরিজিনোসর[৪৭], ড্রোমিওসরিড[৪৮] এবং পাখি[৩৯])। সরিস্কিয়াদের অন্তর্গত ছিল থেরোপড (বাধ্যতামূলকভাবে দ্বিপদী আর বিভিন্ন খাদ্যাভ্যাসযুক্ত) এবং সরোপডোমর্ফ (লম্বা গলাবিশিষ্ট শাকাহারী যাদের অনেকে ছিল অতি-বিবর্তিত চতুষ্পদ)[৪৯][৫০]।
অন্যদিকে অর্নিথিস্কিয়া বা "পাখির-মত-কোমরবিশিষ্ট" (গ্রিক ὀρνίθειος/"অর্নিথেইয়স" বা পাখির এবং ἰσχίον/"ইস্কিয়ন" বা পশ্চাদ্দেশ) ডাইনোসরদের শ্রোণীচক্র আপাতদৃষ্টিতে পাখিদের শ্রোণীচক্রের অনুরূপ ছিল; এদের পিউবিস হাড় আগাগোড়াই পিছনের দিকে বাঁকানো ছিল। কিন্তু পাখিদের থেকে অর্নিথিস্কিয়ান পিউবিসের মূল পার্থক্য ছিল এই যে এদের পিউবিসে একটা সম্মুখবর্তী উপবৃদ্ধিও থাকত যা পাখিদের থাকে না। অর্নিথিস্কিয়াদের বেশ কয়েকটা বিভাগ ছিল এবং এরা সম্ভবত প্রত্যেকেই শাকাহারী ছিল। (বিশেষ দ্রষ্টব্যː পাখির মত কোমর আর গিরগিটির মত কোমর নামদু'টো কিন্তু বিভ্রান্তিকর। পাখিরা গিরগিটির মত কোমরওয়ালা সরিস্কিয়াদের থেকেই বিবর্তিত হয়েছিল)[৪৪]।
-
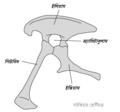 সরিস্কিয়া শ্রোণীচক্রের গঠন (বাঁ দিক)
সরিস্কিয়া শ্রোণীচক্রের গঠন (বাঁ দিক) -
 টির্যানোসরাসের শ্রোণীচক্র (সরিস্কিয়ার উদাহরণ – বাঁ দিক)
টির্যানোসরাসের শ্রোণীচক্র (সরিস্কিয়ার উদাহরণ – বাঁ দিক) -
 অর্নিথিস্কিয়া শ্রোণীচক্রের গঠন (বাঁ দিক)
অর্নিথিস্কিয়া শ্রোণীচক্রের গঠন (বাঁ দিক) -
 এডমন্টোসরাসের শ্রোণীচক্র (অর্নিথিস্কিয়ার উদাহরণ – বাঁ দিক)
এডমন্টোসরাসের শ্রোণীচক্র (অর্নিথিস্কিয়ার উদাহরণ – বাঁ দিক)




শ্রেণিবিন্যাসবিদ্যা
সম্পাদনাবিজ্ঞানী টমাস হল্ৎজ্ ২০০৮ খ্রিষ্টাব্দে মেসোজোয়িক ডাইনোসর প্রজাতিদের একটি তালিকা প্রস্তুত করেন[১]। নিচে সেই তালিকার ভিত্তিতে বিভিন্ন ডাইনোসরের বিবর্তনীয় আন্তঃসম্পর্ক বা শ্রেণিবিন্যাসের একটি সরলীকৃত ছক দেওয়া হল। আরও বিস্তৃত ও সাম্প্রতিক তালিকার জন্য দেখুন ডাইনোসরের শ্রেণিবিন্যাস। বর্তমানে বিলুপ্ত বিভাগগুলোর নামের পাশে ছুরি (†) চিহ্ন বসানো আছে।
- ডাইনোসরিয়া
- সরিস্কিয়া ("গিরগিটির মত পশ্চাদ্দেশ"; এদের অন্তর্গত থেরোপোডা ও সরোপডোমর্ফা)
- থেরোপোডা (প্রত্যেকে দ্বিপদ; অধিকাংশই মাংসাশী)
- †হেরেরাসরিয়া (প্রাথমিক দ্বিপদ মাংসাশী)
- †সিলোফাইসয়ডিয়া (খুদে প্রাথমিক থেরোপড; এদের অন্তর্গত সিলোফাইসিস ও তাদের নিকট জ্ঞাতিরা)
- †ডাইলোফোসরিডি (প্রাথমিক ঝুঁটিওয়ালা মাংসাশী থেরোপড)
- †সেরাটোসরিয়া (সাধারণত বিচিত্র শিংযুক্ত; এরা ছিল ক্রিটেশিয়াসে ভারতীয় উপমহাদেশ সমেত দক্ষিণ গোলার্ধের প্রধান থেরোপড)
- টেটানিউরা (নামের মানে "শক্ত লেজ"; অধিকাংশ থেরোপড এদের অন্তর্ভুক্ত)
- †মেগালোসরয়ডিয়া (বিশাল মাংসাশীদের একটা প্রাথমিক দল; সাঁতারে পটু স্পিনোসরিডরাও এদের অন্তর্গত)
- †কার্নোসরিয়া (অ্যালোসরাস, কার্কারোডন্টোসরাস এবং নিকট জ্ঞাতি)
- সিলুরোসরিয়া (পালকওয়ালা থেরোপড; স্বভাব এবং আয়তন বহু-বিচিত্র)
- †কম্পসোগ্ন্যাথিডি (খুদে হাতওয়ালা সাধারণ প্রাথমিক সিলুরোসর)
- †টির্যানোসরিডি (টির্যানোসরাস এবং নিকট জ্ঞাতি; এদের হাতও ছিল খুদে)
- †অর্নিথোমিমোসরিয়া ("উটপাখির অনুকরণ"; অধিকাংশ দাঁতবিহীন, মাংসাশী থেকে সম্ভাব্য শাকাহারী)
- †আলভারেজ্সরয়ডিয়া (দুই খুদে হাতে একটা করে বড় ধারালো নখওয়ালা ছোট আয়তনের পতঙ্গভুক থেরোপড)
- ম্যানির্যাপ্টোরা ("যারা হাত দিয়ে ছিনতাই করে"; বলা বাহুল্য এদের হাত লম্বা ও নমনীয় আঙুলবিশিষ্ট হত)
- †থেরিজিনোসরিয়া (হাতে বড় থাবা ও ছোট মাথাওয়ালা শাকাহারী থেরোপড)
- †ওভির্যাপ্টরোসরিয়া (অধিকাংশ দাঁতবিহীন; এদের খাদ্যাভ্যাস এবং জীবনযাত্রা সম্বন্ধে নিশ্চিত হওয়া যায়নি)
- †আর্কিওপ্টেরিজিডি (ছোট, ডানাওয়ালা থেরোপড বা প্রাথমিক পাখি)
- †ডাইনোনিকোসরিয়া (ছোট থেকে মাঝারি আয়তন ও পাখির মত আকার; পিছনের পায়ের একটা নখ খুব বড় হত)
- এভিয়ালা (আধুনিক পাখি এবং তাদের বিলুপ্ত জ্ঞাতিসমূহ)
- †স্ক্যান্সোরিওপ্টেরিজিডি (হাতে পরিবর্ধিত তৃতীয় আঙুলযুক্ত ছোট প্রাথমিক এভিয়ালা)
- †অমনিভোরোপ্টেরিজিডি (বেঁটে লেজবিশিষ্ট বড় আয়তনের প্রাথমিক এভিয়ালা)
- †কনফুশিয়াসর্নিথিডি (ছোট দাঁতবিহীন এভিয়ালা)
- †এনান্টিঅর্নিথিস (আদিম উড়ুক্কু গেছো এভিয়ালা)
- ইউঅর্নিথিস (উন্নত উড়ুক্কু পাখি)
- †ইয়ানর্নিথিফর্মিস (ক্রিটেশিয়াস যুগের দাঁতওয়ালা চীনা পাখি)
- †হেস্পারর্নিথিস (বিশেষভাবে অভিযোজিত জলের পাখি)
- এভিস (আধুনিক চঞ্চুবিশিষ্ট পাখি এবং তাদের বিলুপ্ত জ্ঞাতিসমূহ)

- †সরোপডোমর্ফা (ছোট মাথা, লম্বা গলা ও লম্বা লেজযুক্ত শাকাহারী)
- †গুয়াইবাসরিডি (আদিম, ক্ষুদ্রায়তন, সর্বভুক সরোপডোমর্ফ)
- †প্ল্যাটিওসরিডি (বাধ্যতামূলকভাবে দ্বিপদী আদিম "প্রোসরোপড")
- †রিওজাসরিডি (খুদে আদিম সরোপডোমর্ফ)
- †ম্যাসোস্পণ্ডাইলিডি (খুদে আদিম সরোপডোমর্ফ)
- †সরোপোডা (বিশাল এবং ভারী, সাধারণত ১৫ মিটার (৪৯ ফুট)এর বেশি লম্বা; চতুষ্পদ)
- †ভাল্কানোডন্টিডি (থামের মত পা-ওয়ালা প্রাথমিক সরোপড)
- †ইউসরোপোডা ("প্রকৃত" সরোপড)
- †সিটিওসরিডি ("তিমি-সদৃশ সরীসৃপ")
- †তুরিয়াসরিয়া (ইউরোপের জুরাসিক ও ক্রিটেশিয়াস যুগের সরোপডগোষ্ঠী)
- †নিওসরোপোডা ("নতুন সরোপড")
- †ডিপ্লোডোকয়ডিয়া (মাথার খুলি এবং লেজ দীর্ঘায়িত; দাঁত সাধারণত সরু পেন্সিলের আকারের)
- †ম্যাক্রোনারিয়া (বাক্সের মত খুলি; চামচ বা পেন্সিলের মত দাঁত)
- †ব্র্যাকিওসরিডি (লম্বা গলা ও লম্বা অগ্রপদযুক্ত ম্যাক্রোনারিয়ান)
- †টাইটানোসরিয়া (নানা প্রকার; সাধারণত চওড়া কোমরওয়ালা শক্তপোক্ত গড়ন; ক্রিটেশিয়াস যুগে দক্ষিণের মহাদেশগুলোয় সবচেয়ে বেশি দেখা যেত)

- †অর্নিথিস্কিয়া ("পাখির মত কোমরযুক্ত"; নানা রকম দ্বিপদী ও চতুষ্পদ শাকাহারী)
- †হেটারোডন্টোসরিডি (স্পষ্ট শ্বদন্ত যুক্ত খুদে প্রাথমিক অর্নিথিস্কিয়া; শাকাহারী বা সর্বভুক)
- †থাইরিওফোরা (বর্মধারী ডাইনোসর; অধিকাংশই চতুষ্পদ)
- †অ্যাঙ্কিলোসরিয়া (খুব শক্ত আঁশের বর্ম; কারো কারো লেজ হত মুগুর-সদৃশ)
- †স্টেগোসরিয়া (কাঁটা আর পাতের বর্ম)
- †নিঅর্নিথিস্কিয়া ("নতুন অর্নিথিস্কিয়া")
- †অর্নিথোপোডা (আয়তন বিভিন্ন; চতুষ্পদ বা দ্বিপদ; বহুসংখ্যক দাঁত ও নমনীয় চোয়ালের দ্বারা চিবানোর ক্ষেত্রে বাড়তি সুবিধে পেয়েছিল)
- †মার্জিনোসেফালিয়া (মাথার খুলিতে বিশেষ অভিযোজন)
- †প্যাকেসেফালোসরিয়া (খুলিতে গম্বুজ-সদৃশ উপবৃদ্ধিযুক্ত দ্বিপদী)
- †সেরাটোপ্সিয়া (উন্নত শিরস্ত্রাণযুক্ত চতুষ্পদ; অনেকের মাথায় এক বা একাধিক শিংও থাকত)
জীববিদ্যা
সম্পাদনাডাইনোসর সম্বন্ধিত জ্ঞান আহরণের জন্য যে সমস্ত উপাদানের সাহায্য নেওয়া হয় সে'গুলো হল জীবাশ্ম এবং জীবাশ্মীভূত হাড়, অন্যান্য যোগ কলা, বিষ্ঠা, গ্যাস্ট্রোলিথ, পালক, প্রস্তরীভূত পদচিহ্ন, দেহাভ্যন্তরস্থ অঙ্গ, চামড়ার ছাপ ইত্যাদি[৫১][৫২]। বেশ কিছু আলাদা আলাদা শাস্ত্র ডাইনোসর সম্বন্ধে আমাদের ধারণাকে স্পষ্টতর করেছে, যেমন পদার্থবিদ্যা (বিশেষত জীববলবিদ্যা বা বায়োমেকানিক্স, যা ডাইনোসরদের ভর, গমন ও রক্তসংবহন বুঝতে সহায়ক), রসায়ন, জীববিজ্ঞান এবং ভূবিজ্ঞান (পুরাজীববিদ্যা যার একটি শাখা)[৫৩][৫৪]। সম্ভবত ডাইনোসরদের সম্বন্ধে জনমানসে সবচেয়ে বেশি আগ্রহের বিষয় হল তাদের আয়তন এবং আচরণ[৫৫]।
আয়তন
সম্পাদনা
বর্তমান প্রমাণ থেকে বোঝা যায় যে ট্রায়াসিক থেকে জুরাসিক থেকে ক্রিটেশিয়াস যুগে ক্রমশ ডাইনোসরদের গড় আয়তন পরিবর্তিত হয়েছিল[৩২]। দেহের ওজনের ঊর্দ্ধক্রমে সাজালে বেশির ভাগ শিকারী থেরোপড ডাইনোসর ১০০ থেকে ১০০০ কিলোগ্রাম (২২০ থেকে ২২০০ পাউণ্ড) সীমার মধ্যে পড়ত, যেখানে বর্তমান শিকারী স্তন্যপায়ীরা ঐ একই মাপকাঠিতে মাত্র ১০ থেকে ১০০ কিলোগ্রাম (২২ থেকে ২২০ পাউণ্ড) সীমায় পড়ে[৫৬]। সামগ্রিকভাবে মেসোজোয়িক ডাইনোসরদের দেহের গড় ভর ১ থেকে ১০ মেট্রিক টনের মধ্যে থাকত[৫৭]। এই মাপ সিনোজোয়িক স্তন্যপায়ীদের থেকে অনেক বেশি; ন্যাশনাল মিউজিয়াম অফ ন্যাচারাল হিস্ট্রির হিসেব অনুযায়ী স্তন্যপায়ীরা এখন গড় ভরের বিচারে ২ থেকে ৫ কিলোগ্রাম (৫ থেকে ১০ পাউণ্ড) সীমার মধ্যে আছে[৫৮]।
সরোপডরা ছিল বৃহত্তম ও সবচেয়ে ভারী ডাইনোসর। ডাইনোসরদের গোটা রাজত্বকালে ক্ষুদ্রতম সরোপডরাও তাদের বাসস্থানের অন্য যে কোনো প্রাণীর চেয়ে বড় হত, আর বৃহত্তম সরোপডরা ছিল বিশ্বের ইতিহাসে সর্বাপেক্ষা বৃহৎ স্থলচর প্রাণী। ইন্দ্রিকোথেরিয়াম (সর্বকালের বৃহত্তম স্থলচর স্তন্যপায়ী) প্রভৃতি দৈত্যাকার প্রাগৈতিহাসিক স্তন্যপায়ীও সরোপডদের পাশে আয়তনের বিচারে ছিল বামনতুল্য, আর কেবলমাত্র আধুনিক তিমিরা আয়তনে তাদের সাথে পাল্লা দিতে পারে বা তাদের থেকেও বড় হয়[৫৯]। সরোপডদের বিশাল আয়তনের অনেক সম্ভাব্য সুবিধে ছিল, যথা- শিকারী প্রাণীদের থেকে আত্মরক্ষা, শক্তির ব্যয় হ্রাস, দীর্ঘ আয়ু ইত্যাদি; তবে সবচেয়ে গুরুত্বপূর্ণ সুবিধে সম্ভবত হয়েছিল তাদের খাদ্যাভ্যাসের ক্ষেত্রে। বড় প্রাণীদের পৌষ্টিকতন্ত্র ছোট প্রাণীদের পৌষ্টিকতন্ত্র অপেক্ষা উৎকৃষ্টতর হয়, কারণ তাদের পৌষ্টিকনালীতে খাদ্যবস্তু বেশি সময় ধরে থাকে। এর ফলে তারা অপেক্ষাকৃত কম পুষ্টিগুণসম্পন্ন খাবার থেকেও ছোট প্রাণীদের চেয়ে বেশি শক্তি নিংড়ে নিতে পারে। সরোপডদের বিচরণক্ষেত্র ছিল শুষ্ক বা ঋতুবিশেষে শুষ্ক জলবায়ুর অঞ্চল, আর আন্দাজ করা হয় ওই ধরনের জলবায়ুতে উচ্চক্ষমতার পৌষ্টিকতন্ত্র তথা দেহের বিরাট আয়তন তাদের সহায়ক হয়েছিল[৬]।
বৃহত্তম ও ক্ষুদ্রতম
সম্পাদনাবিজ্ঞানীদের পক্ষে সম্ভবত কখনওই সর্বকালের সর্ববৃহৎ ডাইনোসর তথা প্রাণীর পরিচয় সম্পর্কে নিশ্চিত করে কিছু বলা সম্ভব হবে না, কারণ সমস্ত মৃত প্রাণীর খুব অল্প কিছু নমুনাই জীবাশ্মীভূত হয়, আর সমস্ত জীবাশ্মের অধিকাংশই মাটিতে চাপা পড়ে আছে। আবিষ্কৃত নমুনাগুলোর খুব অল্পই গোটা দেহের কঙ্কাল, আর চামড়া ও দেহের অন্যান্য নরম অংশের জীবাশ্ম নমুনা বিরল। অঙ্গসংস্থান ও শারীরবৃত্তীয় ক্রিয়াকলাপের দিক দিয়ে সম্ভাব্য সমতুল্য প্রাণীদের সাথে তুলনা করে একটা গোটা কঙ্কালের ধারণা পাওয়া মুশকিল, আর সেই কঙ্কালের উপর পেশি ও অন্যান্য অঙ্গের অবস্থান নির্দেশ করা খুব বেশি হলে বুদ্ধিদীপ্ত আন্দাজের কাজ মাত্র[৬০]।

যে সমস্ত ডাইনোসরের সম্পূর্ণ কঙ্কাল পাওয়া গেছে তাদের মধ্যে সবচেয়ে বড় ও সবচেয়ে ভারী হল জিরাফাটাইটান ব্রাঙ্কাই। এদের আগে ব্র্যাকিওসরাস -এর উপপ্রজাতি মনে করা হত। ১৯০৭ থেকে ১৯১২ খ্রিষ্টাব্দের মধ্যে তাঞ্জানিয়া থেকে এদের জীবাশ্ম উদ্ধার করা হয়। জার্মানির হাম্বোল্ট মিউজিয়ামে বর্তমানে রক্ষিত কঙ্কালটির বিভিন্ন হাড় প্রকৃতপক্ষে সম আয়তনের একাধিক নমুনা থেকে সংগ্রহ করা[৬১]। এই কঙ্কালটি ১২ মিটার (৩৯ ফু) উঁচু এবং ২২.৫ মিটার (৭৪ ফু) দীর্ঘ, আর সমগ্র প্রাণীদেহটির ভর হওয়ার কথা ৩০০০০ থেকে ৬০০০০ কিলোগ্রাম (৭০০০০ থেকে ১৩০০০০ পাউণ্ড)। আজ পর্যন্ত প্রাপ্ত একক, সম্পূর্ণ ও দীর্ঘতম কঙ্কাল ডিপ্লোডোকাসের ; এটি ২৭ মিটার দীর্ঘ এবং পাওয়া গিয়েছিল মার্কিন যুক্তরাষ্ট্রের ওয়াইওমিং প্রদেশে। ১৯০৭ খ্রিঃ থেকে এটি রাখা আছে ওই দেশেরই কার্নেগী ন্যাচারাল হিস্ট্রি মিউজিয়াম-এ[৬২]।

এর থেকেও বড় ডাইনোসর ছিল, কিন্তু তাদের সম্বন্ধে জানা যায় কেবল কিছু অসম্পূর্ণ কঙ্কাল বা বিক্ষিপ্ত কিছু হাড় থেকে। বড় আয়তনের শাকাহারীদের অধিকাংশ নমুনা আবিষ্কৃত হয়েছে ১৯৭০ এর পর। এদের মধ্যে পড়ে আর্জেন্টিনোসরাস, যাদের ভর ছিল সম্ভবত ৮০০০০ থেকে ১০০০০০ কিলোগ্রাম; আর দীর্ঘতমদের মধ্যে ডিপ্লোডোকাস ছাড়াও আছে সুপারসরাস, যারা ৩৩ মিটার (১০৮ ফু) দীর্ঘ ছিল[৬৩]। উচ্চতম হিসেবে ১৮ মিটার (৫৯ ফু) উঁচু সরোপসাইডন-এর নাম করা যায়, যারা একটা ছ'তলা বাড়ির উপরের তলায় উঁকি দিতে পারত। অবশ্য সমস্ত ডাইনোসরদের মধ্যে সবচেয়ে ভারী আর সবচেয়ে দীর্ঘ প্রজাতি ছিল হয়তো অ্যাম্ফিওসিলিয়াস ফ্র্যাজিলিমাস। দুর্ভাগ্যজনকভাবে এই প্রজাতিটার মেরুদণ্ডের মাত্র একখানা ভাঙা কশেরুকা উদ্ধার করা গিয়েছিল ১৮৭৮ খ্রিষ্টাব্দে, কিন্তু এখন সেটাও হারিয়ে গেছে। ঐ একটা হাড়ের ছবি দেখে অঙ্ক কষে বলা যায় গোটা প্রাণীটার দৈর্ঘ্য ছিল সম্ভবত ৫৮ মিটার (১৯০ ফু) আর ভর ১২০০০০ কিগ্রাঃ (২৬০০০০ পাউণ্ড)[৬]। আবিষ্কৃত বৃহত্তম মাংসাশী ডাইনোসর হল স্পিনোসরাস, ১৬ থেকে ১৮ মিটার (৫২ থেকে ৬০ ফুট) দীর্ঘ আর ৮১৫০ কিলোগ্রাম ভারী[৬৪]। অন্যান্য বড় মাপের মাংসাশীরা হল জাইগ্যান্টোসরাস, কার্কারোডন্টোসরাস, টির্যানোসরাস ইত্যাদি[৬৫]।
উড়তে অক্ষম ডাইনোসরদের মধ্যে ক্ষুদ্রতম প্রজাতিগুলোর আয়তন হত পায়রার মত[৬৬]। স্বাভাবিকভাবেই বোঝা যায় এরা ছিল পাখিদের নিকটতম থেরোপড। যেমন, অ্যাঙ্কিয়র্নিস হাক্সলেয়ি -এর কঙ্কালের মোট দৈর্ঘ্য ৩৫ সেন্টিমিটার (১.১ ফুট)[৬৬][৬৭]। এ. হাক্সলেয়ি হল আজ পর্যন্ত বর্ণিত ক্ষুদ্রতম পূর্ণবয়স্ক উড়তে অক্ষম ডাইনোসর। এদের ভর কমবেশি ১১০ গ্রামের কাছাকাছি থাকত[৬৭]। ক্ষুদ্রতম শাকাহারী উড়তে অক্ষম ডাইনোসরদের মধ্যে ছিল মাইক্রোসেরাটাস এবং ওয়ানানোসরাস ; উভয়েরই দৈর্ঘ্য প্রায় ৬০ সেমি (২.০ ফু)[১][৬৮]।
আচরণ
সম্পাদনা
আধুনিক পখিদের অনেক প্রজাতি সামাজিক জীবনযাপনে অভ্যস্ত এবং তারা বড় দল তৈরি করে বসবাস করে। বিজ্ঞানীরা স্বীকার করেন যে পাখি এবং কুমিরদের (যারা বিবর্তনে পাখিদের নিকটতম জ্ঞাতি) বেশ কিছু আচরণ বিলুপ্ত ডাইনোসর প্রজাতিদের মধ্যেও দেখা যেত। ডাইনোসরদের আচরণ সম্বন্ধে অনুমান করা হয় তাদের কঙ্কালের ভঙ্গী, বাসস্থান, অঙ্গসঞ্চালনের কম্পিউটার অনুকৃতি এবং বর্তমান বাস্তুতন্ত্রের অনুরূপ ধাপে থাকা প্রাণীদের সাথে তুলনার মাধ্যমে[৫৩]।
ডাইনোসরদের মধ্যে দলবদ্ধ জীবনযাত্রার প্রথম প্রমাণ পাওয়া যায় ১৮৭৮ খ্রিঃ আবিষ্কৃত ৩১ টা ইগুয়ানোডন বার্নিসার্টেনসিস-এর জীবাশ্ম থেকে, যদিও প্রথমে মনে করা হয়েছিল এই প্রাণীগুলো দুর্ঘটনাক্রমে বন্যার সময়ে কোনো গভীর গহ্বরের মধ্যে পড়ে গিয়ে একসাথে প্রাণ হারায়[৬৯]। পরবর্তীকালে এ'রকম যৌথ সমাধির আরও নিদর্শন পাওয়া গেছে। এই সমস্ত নিদর্শন এবং একই জায়গায় অনেক ডাইনোসরের জীবাশ্মীভূত পায়ের ছাপের আধিক্য অনেক রকম ডাইনোসরের মধ্যে দলবদ্ধ আচরণের দিকে ইঙ্গিত করে। শত শত, এমনকি হাজার হাজার পায়ের ছাপ প্রমাণ করে হ্যাড্রোসরিড ডাইনোসরেরা আধুনিক আমেরিকান বাইসন বা আফ্রিকান স্প্রিংবক হরিণের মত বিশাল বিশাল দল হিসেবে চলাফেরা করত। সরোপডদের পায়ের ছাপের নিদর্শন প্রমাণ করে এদের একাধিক প্রজাতির সদস্যেরা কখনও কখনও একই দলের অন্তর্ভুক্ত থাকত (যেমন, অধুনা ইংল্যান্ডের অক্সফোর্ডশায়ারে)[৭০], যদিও তাদের দলের গঠন সম্বন্ধে বিশেষ কিছু জানা যায়নি[৭১]। দলবদ্ধ জীবনযাত্রার বিবর্তনের কারণ হতে পারে আত্মরক্ষা, পরিযান অথবা শাবকদের নিরাপত্তা সুনিশ্চিত করা। অল্পবয়স্ক থেরোপড, সরোপড, অ্যাঙ্কিলোসর, অর্নিথোপড এবং সেরাটোপ্সিয়ানেরা যে অনেক ক্ষেত্রে আলাদা দল তৈরি করে থাকত তার প্রমাণ পাওয়া গেছে। উদাহরণ হিসেবে মঙ্গোলিয়ায় প্রাপ্ত ২০ টি ১ থেকে ৭ বছর বয়স্ক সিনর্নিথোমিমাসের জীবাশ্মের কথা বলা যায়। এরা কাদায় আটকা পড়ে মারা গিয়েছিল বলে ধারণা করা হয়[৭২]। কোনো কোনো থেরোপড ডাইনোসরও আধুনিক নেকড়ের মত দল বেঁধে বড় শিকার ধরত বলে অনুমান করা হয়[৭৩][৭৪]। অবশ্য এই ধরনের আচরণ পাখি, কুমির ও অন্যান্য সরীসৃপদের মধ্যে আর দেখা যায় না, আর ডাইনোনিকাস বা অ্যালোসরাস-এর জীবাশ্মের যে সমস্ত নিদর্শনে শিকারের পাশেই একাধিক শিকারী প্রাণীর অবশেষ পাওয়া গেছে সেগুলো শিকারের অধিকার নিয়ে শিকারীদের মধ্যে ঘটে যাওয়া খুনোখুনির ফলও হতে পারে; আধুনিক স্তন্যপায়ীদের মধ্যেও এরকম ঘটে থাকে[৭৫]।

মার্জিনোসেফালিয়া, ল্যাম্বিওসরিন ও ক্ষেত্রবিশেষে থেরোপড ডাইনোসরদের শিং, ঝুঁটি এবং শিরস্ত্রাণগুলো প্রায়শই কার্যকর আত্মরক্ষার পক্ষে খুব দুর্বল। তাই আন্দাজ করা হয় এগুলো যৌন উদ্দীপনা বা সতর্কীকরণের উদ্দেশ্যেই ব্যবহৃত হত (আধুনিক ময়ূরের পেখম বা বিষধর ব্যাঙ প্রভৃতির উজ্জ্বল বর্ণের অনুরূপ), যদিও ডাইনোসরদের যৌন সঙ্গম এবং এলাকা চিহ্নিতকরণ সম্বন্ধে খুব কমই জানা গেছে। কামড়ের ফলে মাথায় প্রাপ্ত আঘাতের চিহ্ন থেকে বোঝা যায় যে অন্তত থেরোপডেরা এলাকা দখলের জন্য পরস্পরের সাথে আগ্রাসী দ্বন্দ্বে লিপ্ত হত[৭৬]।
আচরণের দিক থেকে ভীষণ গুরুত্বপূর্ণ একটি জীবাশ্ম পাওয়া গিয়েছিল ১৯৭১ খ্রিঃ গোবি মরুভূমি থেকে; একটি প্রোটোসেরাটপ্স কে আক্রমণরত একটি ভেলোসির্যাপ্টর[৭৭]। এ'থেকে ডাইনোসরদের মধ্যে একে অপরকে আক্রমণের প্রত্যক্ষ প্রমাণ মেলে[৭৮]। জীবন্ত শিকারকে আক্রমণের আরও একটা উদাহরণ হল এডমন্টোসরাস-এর একটি জীবাশ্ম, যার লেজে টির্যানোসরাস-এর কামড়ের চিহ্ন রয়েছে। কামড় খেয়েও এডমন্টোসরাস টি কোনোক্রমে বেঁচে গিয়েছিল[৭৮]। এছাড়া মাদাগাস্কার থেকে প্রাপ্ত মাজুঙ্গাসরাস প্রভৃতি কোনো কোনো ডাইনোসর প্রজাতির মধ্যে ক্যানিবালিজ্ম্ বা নিজের প্রজাতিরই অন্যান্য সদস্যকে ভক্ষণ করার প্রবৃত্তির প্রমাণ পাওয়া গেছে[৭৯]।
ডাইনোসর এবং আধুনিক পাখিদের চোখের শ্বেতমণ্ডলীয় বলয় তুলনা করে ডাইনোসরদের দৈনিক সক্রিয়তার সময় নির্ধারণের চেষ্টা করা হয়। অধিকাংশ ডাইনোসর দিবাচর ছিল বলে ধারণা করা হলেও কিছু কিছু ড্রোমিওসরিড, জুরাভেনাটর এবং মেগাপ্নোসরাস প্রভৃতি ছোট আয়তনের মাংসাশীরা খুব সম্ভবত নিশাচর ছিল। সেরাটোপ্সিয়ান, সরোপডোমর্ফ, হ্যাড্রোসরিড এবং অর্নিথোমিমোসর প্রভৃতি বড় থেকে মাঝারি আয়তনের শাকাহারীরা দিনের কোনো কোনো সময় কিছুক্ষণের জন্য সক্রিয় হত, যদিও ছোট মাপের অর্নিথিস্কিয়ান অ্যাজিলিসরাস দিবাচর ছিল মনে করা হয়[৮০]।
প্রাপ্ত জীবাশ্ম প্রমাণের উপর নির্ভর করে অরিক্টোড্রোমিয়াস ইত্যাদি কিছু খুদে অর্নিথিস্কিয়ান প্রজাতির মধ্যে আংশিকভাবে মাটিতে গর্ত খুঁড়ে বসবাসের প্রমাণ মেলে[৮১]। বর্তমানে বেশ কিছু পাখি গাছে চড়তে পারে, আর মেসোজোয়িক মহাযুগের পাখিরাও ক্ষেত্রবিশেষে এই কাজে দড় ছিল, যেমন এনান্টিঅর্নিথিন[৮২]। অবশ্য অল্প কিছু প্রজাতি ছাড়া বেশিরভাগ ডাইনোসর স্থল গমনেই অভ্যস্ত ছিল। মাটির উপর ডাইনোসরেরা কী রকম ভাবে চলাফেরা করত তার একটা স্পষ্ট ধারণা থাকলে তাদের আচরণের নানা খুঁটিনাটি বোঝার খুব সুবিধে হয়। বিশেষত জীববলবিদ্যা বা বায়োমেকানিক্স শাস্ত্রটি এই ক্ষেত্রে গুরুত্বপূর্ণ বেশ কিছু পথের সন্ধান দিয়েছে। যেমন, পেশি ও অভিকর্ষের দ্বারা ডাইনোসরদের কঙ্কালের উপর প্রযুক্ত বলের তীব্রতা ও অভিমুখ পর্যালোচনা করে জানা যায় ডাইনোসরেরা কত জোর দৌড়তে পারত[৮৩], ডিপ্লোডোকয়েডরা সত্যিই তাদের চাবুকের মত লেজের ঝাপটায় সনিক বুম তৈরি করতে পারত কি না[৮৪], আর সরোপডেরা জলে ভাসতে পারত কি না[৮৫]।
ভাব বিনিময়
সম্পাদনা
আধুনিক পাখিরা প্রধানত নানা রকম দৃশ্যমান ও শ্রবণযোগ্য সংকেতের মাধ্যমে পরস্পরের সাথে ভাব বিনিময়ের জন্য বিখ্যাত, আর জীবাশ্মীভুত ডাইনোসরদের মধ্যে প্রদর্শন-অঙ্গের নিদর্শনের বৈচিত্র্য ও প্রাচুর্য দেখে মনে করা হয় ভাব বিনিময়ের বিষয়টি ডাইনোসরদের মধ্যেও গুরুত্বপূর্ণ ছিল। অবশ্য ডাইনোসরদের ডাক কেমন হত ও কী কাজে ব্যবহৃত হত তা নিয়ে নিশ্চিত হওয়া যায় না। ২০০৮ খ্রিঃ পুরাজীববিদ ফিল সেন্টার ডাইনোসর সমেত অন্যান্য মেসোজোয়িক প্রাণীর দেহে শব্দ করার ক্ষমতার নিদর্শন সংক্রান্ত একটি পরীক্ষা চালান[৮৬]। সেন্টার বুঝতে পারেন চলচ্চিত্র ও তথ্যচিত্রে হামেশা ডাইনোসরদের হুঙ্কার দিতে দেখা গেলেও মেসোজোয়িক ডাইনোসরদের অধিকাংশই খুব সম্ভবত কোনও রকম শব্দ করতে পারত না, যদিও ল্যাম্বিওসরিনদের মাথায় অবস্থিত ফাঁপা নলাকার ঝুঁটিগুলো বিভিন্ন রকম শব্দ সৃষ্টি করার সময় অনুনাদ প্রকোষ্ঠ হিসেবে ব্যবহৃত হয়ে থাকার সম্ভাবনা আছে[৮৭][৮৮]। এই সিদ্ধান্তে উপনীত হওয়ার আগে সেন্টার আধুনিক সরীসৃপ ও পাখিদের দেহে শব্দসৃষ্টিকারী অঙ্গসমূহের বিন্যাস লক্ষ্য করেন। তিনি দেখেন, সরীসৃপদের মধ্যে একাধিকবার স্বরযন্ত্রের (ইংরেজিতে 'Larynx') বিবর্তন হয়েছে। কুমিরেরাও এর ব্যতিক্রম নয়, যে কারণে তারা এক প্রকার ঘড়ঘড়ে আওয়াজ করতে পারে। কিন্তু পাখিদের কোনো স্বরযন্ত্র নেই। পাখির ডাকের শব্দ উৎপন্ন হয় সিরিংক্স নামক একটি অঙ্গ থেকে, যেটি সমস্ত পাখির দেহেই বর্তমান এবং যেটির সাথে স্বরযন্ত্রের কোনও সম্পর্ক নেই। এ'থেকে বোঝা যায় অন্যান্য সরীসৃপের স্বরযন্ত্রের থেকে সম্পূর্ণ আলাদাভাবে সিরিংক্সের বিবর্তন হয়েছিল। সিরিংক্সকে ঠিকঠাক কাজ করতে হলে বায়ুথলির প্রয়োজন হয়; বিশেষত কন্ঠাস্থির কাছে অবস্থিত "কন্ঠাস্থি বায়ুথলি" এর কাজে অপরিহার্য। এই বায়ুথলিটি সংলগ্ন হাড়গুলোয় বিশেষ প্রকার দাগ বা ফুটোর সৃষ্টি করে। বিশেষত প্রগণ্ডাস্থি বা হিউমেরাস হাড়ে বেশ বড় একটা ফুটো এর অবস্থানের ফলে তৈরি হয়। সরিস্কিয়ান ডাইনোসরদের দেহে বায়ুথলির অবস্থান ও নানা রকম বিন্যাস দেখা গেলেও ডাকের জন্য প্রয়োজনীয় কন্ঠাস্থি বায়ুথলির নিদর্শন এনান্টিঅর্নিথিন-এর আগে কোনও জীবাশ্মে পাওয়া যায় না। এর একমাত্র ব্যতিক্রম ছিল এরোস্টিয়ন, যারা সম্ভবত শব্দ করা ছাড়া অন্য কোনও কারণে পাখিদের থেকে আলাদাভাবে কন্ঠাস্থি বায়ুথলির উদ্ভব ঘটিয়েছিল[৮৬]।
অতএব শব্দ সৃষ্টি করতে সক্ষম সিরিংক্সের নিদর্শনযুক্ত প্রথম ডাইনোসর হল এনান্টিঅর্নিথিন পাখিরা। এদের চেয়ে প্রাচীন পক্ষীগোত্রীয় আর্কোসরেরা সম্ভবত গলা থেকে কোনও আওয়াজ করতে পারত না। কিন্তু জীবাশ্ম থেকে প্রাপ্ত প্রচুর নিদর্শন থেকে জানা যায় প্রাথমিক ডাইনোসরেরা ভাব বিনিময়ের ক্ষেত্রে দৃশ্যমান সঙ্কেতের বহুল ব্যবহার করত। তাদের অজস্র প্রদর্শন-অঙ্গের মধ্যে পড়ে বিশেষ আকারের শিং, শিরস্ত্রাণ, ঝুঁটি, পাতের আকারের উপবৃদ্ধি এবং পালক। আন্দাজ করা হয় এই অঙ্গগুলোর রঙও হয়তো ছিল উজ্জ্বল ও বিচিত্র, এবং সামগ্রিকভাবে এই ব্যবস্থাটি আজকের টিকটিকি-গিরগিটি-গোসাপদের কথা মনে পড়িয়ে দেয়। ডাইনোসরদের মতোই এদেরও অধিকাংশ প্রজাতি বোবা (যদিও শোনার ক্ষমতা প্রত্যেকেরই খুব ভাল), আর ভাব আদানপ্রদানের জন্য এরাও নানা রকম রঙ ও প্রদর্শন-অঙ্গের সাহায্য নেয়[৮৬]।
এছাড়া ডাইনোসরেরা অন্য উপায় অবলম্বন করেও শব্দ সৃষ্টি করত। কয়েক প্রকার সরীসৃপ ও অন্যান্য প্রাণী ডাক ছাড়া অন্যভাবে শব্দ করতে পারে, যেমন সাপের হিস্, চোয়ালে চোয়ালে ঘর্ষণ, জল ছিটিয়ে শব্দ করা, ডানা ঝাপটানোর আওয়াজ প্রভৃতি। ডাইনোসরেরাও এই পদ্ধতিগুলো জানত বলে অনুমান করা হয়[৮৬]।
প্রজনন
সম্পাদনা
সমস্ত ডাইনোসরই মূলত ক্যালশিয়াম কার্বনেট নির্মিত শক্ত খোলায় ঢাকা অ্যামনিওটিক ডিম থেকে জন্ম নেয়[৮৯]। ডিম পাড়ার জায়গা হল বিশেষভাবে নির্মিত বাসা। পাখিদের অনেক প্রজাতি বিচিত্র আকারের বাসা তৈরি করেː- পেয়ালা, গম্বুজ, প্লেট, খড়কুটোর গদি, ঢিপি বা সুড়ঙ্গের মত দেখতে বাসার অস্তিত্ব আছে[৯০]। অবশ্য কিছু কিছু পাখি বাসা বানায় না; 'মুরে' পাখি প্রভৃতি কেউ কেউ যারা খাড়া পাহাড়ের গায়ে বসবাস করে তারা পাথরের উপরেই ডিম রাখে, আর এম্পারার পেঙ্গুইন ডিমে তা দেয় তাদের পেটের তলায় দু'পায়ের ফাঁকে ডিম গুঁজে নিয়ে। প্রাচীন পাখি ও উড়তে অক্ষম ডাইনোসরেরা প্রায়ই খুব বড় গোষ্ঠীগত বাসা বানিয়ে তাতে বহুসংখ্যক ডিম রাখত। ডিমে তা দেওয়ার কাজ ছিল প্রধানত পুরুষদের। আধুনিক পাখিদের কার্যকর ডিম্বনালীর সংখ্যা এক হলেও আদিম পাখি ও উড়তে অক্ষম ডাইনোসরেদের দু'টো করে ডিম্বনালীই সক্রিয় ছিল, যেমন আজকের কুমিরের দেহে দেখা যায়। ট্রুডন জাতীয় কিছু কিছু উড়তে অক্ষম ডাইনোসর কয়েক খেপে ডিম পাড়ত, অর্থাৎ মা ডাইনোসর দু'একদিন অন্তর দু'একটা করে ডিম পাড়ত, কিন্তু সমস্ত ডিম পাড়া হয়ে যাওয়ার পর একসাথে তা দিতে শুরু করত যাতে সবকটা ডিম একই সাথে ফুটে বাচ্চা হয়[৯১]।
ডিম পাড়ার সময় স্ত্রী প্রাণীরা তাদের পায়ের বাইরের দিককার শক্ত হাড় আর অস্থিমজ্জার মাঝখানে ক্যালশিয়াম সমৃদ্ধ বিশেষ একরকম মেডুলারি হাড় তৈরি করে যা ডিমের খোলা নির্মাণে ব্যবহৃত হয়। উড়তে অক্ষম ডাইনোসরদের মধ্যে টির্যানোসরাস রেক্স-এর একটি কঙ্কালে প্রথম এই মেডুলারি হাড়ের অবস্থানের প্রমাণ পাওয়া যায়। এরই ফলে প্রথম বার পুরাজীববিদরা কোনো জীবাশ্ম নমুনার লিঙ্গ নির্ণয়ে সক্ষম হন। পরবর্তী গবেষণায় অন্যতম কার্নোসর অ্যালোসরাস এবং অন্যতম অর্নিথোপড টেনন্টোসরাসের জীবাশ্মে মেডুলারি হাড়ের সন্ধান মিলেছে। যেহেতু টেনন্টোসরাসের বিবর্তনের ধারাটি অ্যালোসরাস ও টির্যানোসরাসের থেকে ডাইনোসর বিবর্তনের একেবারে প্রাথমিক পর্যায়েই আলাদা হয়ে গিয়েছিল, তাই বোঝা যায় মেডুলারি হাড়ের অস্তিত্ব হল সমস্ত ডাইনোসরের একটি সাধারণ বৈশিষ্ট্য[৯২]।

আধুনিক পাখিদের আরো একটা বৈশিষ্ট্য হল বাচ্চা জন্মানোর পর তার লালনপালন। মন্টানায় ১৯৭৮ খ্রিঃ জ্যাক হর্নার, মাইয়াসরা ("ভালো-মা-গিরগিটি")-এর একটি বাসা আবিষ্কার করেন এবং সেখান থেকেই অর্নিথোপড ডাইনোসরদের মধ্যে শিশু জন্মের অনেকদিন পর পর্যন্ত তাদের লালনপালনের প্রমাণ পাওয়া যায়। এ'থেকে আন্দাজ করা হয় হয়তো অন্যান্য ডাইনোসরদের মধ্যেও আচরণের এই বৈশিষ্ট্যটি দেখা যেত[৯৩]। প্রমাণ আছে যে অন্যান্য অ-থেরোপড ডাইনোসর, যেমন প্যাটাগোনিয়ার টাইটানোসর জাতীয় সরোপডরাও বড় বাসা তৈরি করে থাকত (আবিষ্কার ১৯৯৭ খ্রিঃ)[৯৪]। ১৯৯৩ খ্রিঃ মঙ্গোলিয়ায় একটি ওভির্যাপ্টরিড ডাইনোসর চিতিপতি অস্মোল্স্কি-এর জীবাশ্ম পাওয়া যায়, যেটি একটি বাসার উপর ডিমে তা দেওয়ার ভঙ্গিতে বসে ছিল[৯৫]। এ'থেকে ইঙ্গিত পাওয়া যায় এদের দেহে উষ্ণতারোধী পালকের বিবর্তন হয়েছিল যা ডিম গরম রাখার কাজে সহায়ক[৯৬]। সন্তানের লালনপালন যে সমস্ত ডাইনোসরের সাধারণ বৈশিষ্ট্য ছিল তার সপক্ষে আরো প্রমাণ আছে। যেমন, অন্যতম প্রোসরোপড ম্যাসোস্পণ্ডাইলাসের দাঁতবিহীন একটি ভ্রূণের জীবাশ্ম পাওয়া গেছে, যা থেকে বোঝা যায় ঐ প্রজাতির সদ্যোজাত শিশুদের খেতে দেওয়ার জন্য মা বাবার বিশেষ যত্নের দরকার হত[৯৭]। উত্তর-পশ্চিম স্কটল্যান্ডের স্কাই দ্বীপে প্রাপ্ত পদচিহ্নের জীবাশ্ম থেকে অর্নিথোপড ডাইনোসরদের সন্তানপালনের প্রমাণ মেলে[৯৮]। ডাইনোসরদের অধিকাংশ প্রধান গোষ্ঠীরই বাসা ও ডিমের জীবাশ্ম পাওয়া গেছে আর মনে করা হয় তারা প্রত্যেকেই জন্মের আগে বা জন্মের অব্যবহিত পরে তাদের সন্তানদের যত্ন করত[৯৯]।
শারীরবৃত্ত
সম্পাদনা
যেহেতু আধুনিক পাখি আর কুমির উভয়েরই হৃৎপিণ্ড চার প্রকোষ্ঠযুক্ত (কুমিরের ক্ষেত্রে একটু বেশি উন্নত), তাই মনে করা হয় এই ধরনের হৃৎপিণ্ড সমস্ত আর্কোসরের সাধারণ বিবর্তনীয় বৈশিষ্ট্য, যে তালিকায় উড়তে অক্ষম ডাইনোসরেরাও পড়ে[১০০]। যদিও আধুনিক পাখিরা প্রত্যেকেই উচ্চ বিপাক হার যুক্ত উষ্ণশোণিত প্রাণী, কিন্তু বিবর্তনের ধারায় ডাইনোসরেরা ঠিক কখন উষ্ণশোণিত হয়ে উঠেছিল তা নিয়ে বিজ্ঞানীদের মধ্যে ১৯৬০ এর দশক থেকে প্রবল বিতর্ক চলছে। উড়তে অক্ষম ডাইনোসরেরা উষ্ণশোণিত, অনুষ্ণশোণিত নাকি দুই প্রকারেরই হত সে'ব্যাপারে আপাতত কোনো ঐকমত্য্য নেই[১০১]।
উড়তে অক্ষম ডাইনোসরদের আবিষ্কারের পরে পুরাজীববিদরা প্রাথমিকভাবে তাদের অনুষ্ণশোণিত হিসেবে ধারণা করেছিলেন। এ'থেকে সিদ্ধান্ত করা হয়েছিল তারা অপেক্ষাকৃত ধীর ও অলস প্রকৃতির জীব, যদিও আধুনিক সরীসৃপদের অনেকেই দেহের তাপমাত্রা নিয়ন্ত্রণের জন্য বহিঃপরিবেশের উপর নির্ভরশীল হলেও ক্ষিপ্র চালচলনে অভ্যস্ত। যাই হোক, অবলুপ্ত ডাইনোসরদের ঠাণ্ডা রক্তের প্রাণী মনে করার প্রচলন বজায় ছিল ১৯৬৮ খ্রিঃ পর্যন্ত; ঐ বছর বিজ্ঞানী রবার্ট ব্যাকার তাদের উষ্ণশোণিত হিসেবে প্রতিপন্ন করে একটি গুরুত্বপূর্ণ গবেষণাপত্র প্রকাশ করেন[১০২]।
সাম্প্রতিককালে প্রাপ্ত প্রমাণ থেকে বোঝা যায় অন্তত কিছু আদিম উড়তে অক্ষম ডাইনোসর ও পাখি ঠাণ্ডা ও নাতিশীতোষ্ণ জলবায়ু অঞ্চলে বসবাস করত এবং দেহাভ্যন্তরীণ জৈবিক ব্যবস্থার মাধ্যমে নিজেদের দেহের তাপমাত্রাও নিয়ন্ত্রণ করতে পারত (বড় প্রাণীরা বিশাল আয়তনের মাধ্যমে আর ছোট প্রাণীরা পালক বা অনুরূপ কোনো দেহ-আবরকের মাধ্যমে)। মেসোজোয়িক ডাইনোসরদের দেহে উষ্ণশোণিত ব্যবস্থার আরও প্রমাণ পাওয়া গেছে অস্ট্রেলিয়া ও অ্যান্টার্কটিকায় মেরু ডাইনোসর-দের জীবাশ্ম আবিষ্কার হওয়ার পর। কিছু কিছু ডাইনোসর জীবাশ্মের হাড়ের অভ্যন্তরে রক্তবাহী নালিকার যে বিন্যাস দেখা যায়, তা-ও উষ্ণশোণিত প্রাণীরই অনুরূপ। তা সত্ত্বেও ডাইনোসরদের দেহের তাপমাত্রা নিয়ন্ত্রণের যথাযথ ব্যবস্থা ও তার বিবর্তনের মডেলটি সম্পর্কে বিজ্ঞানীদের মধ্যে বিতর্ক চলছে[১০৩]।
সরিস্কিয়ান ডাইনোসরদের দেহে পাখি-সদৃশ শ্বসনতন্ত্রের বিবর্তনের মাধ্যমে বিপাক হারের বৃদ্ধি সম্ভব হয়েছিল। এই ধরনের শ্বসনতন্ত্রের বৈশিষ্ট্য হল একাধিক বায়ুথলির অবস্থান, যার সাহায্যে ফুসফুসের ক্ষমতা অনেকখানি বৃদ্ধি পায় আর কঙ্কালতন্ত্রের অনেকগুলো হাড়ের মধ্যেও বায়ুথলির সম্প্রসারণের ফলে হাড়গুলো ফাঁপা হয়ে যায়[১০৪]। আদিম পক্ষীসদৃশ শ্বসনতন্ত্রের অধিকারী সরীসৃপেরা সম্ভবত অনুরূপ আয়তন ও গঠনের স্তন্যপায়ীদের চেয়েও বেশি সক্রিয় থাকতে পারত। প্রচুর অক্সিজেনের উৎকৃষ্ট সরবরাহ ব্যবস্থার পাশাপাশি এই বায়ুথলিগুলো দেহের অতিরিক্ত তাপ বাইরে বার করে দিতেও সক্ষম ছিল, আর এই ব্যবস্থাটার গুরুত্বও ছিল অপরিসীম, কারণ বিশাল আয়তনের সক্রিয় প্রাণীরা শুধুমাত্র চামড়ার মাধ্যমে তাদের দেহের সমস্ত অতিরিক্ত তাপ পরিবেশে মোচন করতে পারে না[১০৫]।
অন্যান্য সরীসৃপের মত ডাইনোসরেরাও ছিল প্রধানত ইউরিকোটেলিক, অর্থাৎ তাদের বৃক্ক রক্তস্রোত থেকে নাইট্রোজেনঘটিত বর্জ্য পদার্থ নিষ্কাষণ করে তা ইউরিক অ্যাসিড হিসেবে গবিনীর মাধ্যমে অন্ত্রের মধ্যে ত্যাগ করত; ইউরিক অ্যাসিডকে ইউরিয়া বা অ্যামোনিয়ায় পরিণত করত না। অধিকাংশ জীবন্ত ইউরিকোটেলিক প্রাণীদেহ থেকে মলের সাথে ইউরিক অ্যাসিডের মিশ্রণ একটি অর্ধকঠিন বর্জ্য হিসেবে পরিত্যক্ত হয়[১০৬][১০৭][১০৮]। যদিও কোনো কোনো আধুনিক পাখি (যেমন হামিংবার্ড) অন্তত আংশিকভাবে অ্যামোনোটেলিক, অর্থাৎ এরা অধিকাংশ নাইট্রোজেনঘটিত বর্জ্যপদার্থ অ্যামোনিয়া হিসেবে ত্যাগ করে[১০৯]। আবার এরা স্তন্যপায়ীদের মত ক্রিয়েটিনিন-এর বদলে ক্রিয়েটিন ত্যাগ করে। অন্ত্র থেকে বহিষ্কৃত বর্জ্যের সাথেই এই পদার্থটিও অবসারণী ছিদ্রের মাধ্যমে দেহের বাইরে নির্গত হয়[১১০][১১১]। এছাড়া অনেক প্রজাতি হজম না হওয়া খাবার পেলেট হিসেবেও মুখ থেকে উগরে দেয়, আর ডাইনোসরদের থেকে আগত সম্ভাব্য পেলেটের জীবাশ্ম পাওয়া গেছে ক্রিটেশিয়াস যুগের পাথরের স্তরে[১১২]।
পাখির উৎপত্তি
সম্পাদনাডাইনোসরেরা যে পাখিদের পূর্বপুরুষ হতে পারে এ'কথা প্রথম বলেন টমাস হেনরি হাক্সলি, ১৮৬৮ খ্রিষ্টাব্দে[১১৩]। বিংশ শতাব্দীর শুরুতে জেরার্ড হাইলমানের গবেষণার পর এই ধারণার পরিবর্তে ডাইনোসর ও পাখিদেরকে সাধারণ থেকোডন্ট-দের আলাদা আলাদা বংশধর বলে চিহ্নিত করা হয়। এর প্রমাণ হিসেবে ডাইনোসরদের জীবাশ্মে 'কন্ঠাস্থির অনুপস্থিতির' নিদর্শন দেওয়া হত [১১৪]। কিন্তু পরবর্তী অনুসন্ধান প্রমাণ করে যে ডাইনোসরদের কন্ঠাস্থি প্রকৃতপক্ষে অনুপস্থিত নয়; পাখিদের মতোই তাদের দু'টি কন্ঠাস্থি জুড়ে গিয়ে ফারকুলা নামক একক হাড় হিসেবে অবস্থান করে[৩৯]। ওভির্যাপ্টর-এর জীবাশ্মে এর আগে ১৯২৪ খ্রিঃ এই হাড়ের খোঁজ পাওয়া গিয়েছিল, কিন্ত বিজ্ঞানীরা তখন সেটাকে আন্তঃকন্ঠাস্থি মনে করেছিলেন[১১৫]। ১৯৭০ এর দশকে বিজ্ঞানী জন অস্ট্রম ডাইনোসর থেকে পাখির উৎপত্তির মতবাদের পুনঃপ্রতিষ্ঠা করেন[১১৬], এবং পরবর্তীকালে ক্ল্যাডিস্টিক বিশ্লেষণের বহুল প্রচলন ও প্রচুর থেরোপড ও আদিম পাখির জীবাশ্মের আবিষ্কারের[১১৭] সাথে সাথে মতটি অধিক গুরুত্ব পেতে থাকে[১১৮]। এই প্রসঙ্গে বিশেষভাবে উল্লেখযোগ্য য়িক্সিয়ান প্রস্তরক্ষেত্র, যেখান থেকে বহুসংখ্যক আদিম পাখি ও থেরোপডের জীবাশ্ম পাওয়া গেছে, অনেক ক্ষেত্রে কোনো না কোনো পালক সমেত[৩৯]। পাখিদের সাথে থেরোপড ডাইনোসরদের শারীরস্থানিক বৈশিষ্ট্যের মিলের সংখ্যা একশোরও বেশি, আর ঐ ডাইনোসরদেরকেই এখন বিবর্তনের ধারায় পাখিদের নিকটতম জ্ঞাতি বলে মনে করা হয়[১১৯]। সিলুরোসরদের অন্তর্গত ম্যানির্যাপ্টোরা বিভাগটি পাখিদের নিকটতম[৩৯]। অ্যালান ফেদুচ্চিয়া, ল্যারি মার্টিন ও মুষ্টিমেয় অপর কয়েকজন বিজ্ঞানী বিকল্প বিবর্তনীয় ধারার প্রস্তাব করেছেন। এগুলোর মধ্যে পড়ে হাইলমানের প্রাথমিক আর্কোসর মতবাদের পুনর্নবীকরণ এবং ম্যানির্যাপ্টোরাদেরকে পাখিদের পূর্বপুরুষ হিসেবে মেনে নিয়েও তাদের ডাইনোসরদের থেকে আলাদা করে দেখা। এই শেষোক্ত মতবাদে ডাইনোসরদের সাথে ম্যানির্যাপ্টোরার মিলটি অভিসারী বিবর্তনের নমুনা হিসেবে গণ্য করা হয়[২]।
পালকযুক্ত ডাইনোসর
সম্পাদনা
পালক হল আধুনিক পাখিদের অন্যতম বিশিষ্ট শনাক্তকরণ বৈশিষ্ট্য, আর অন্যান্য ডাইনোসরদের মধ্যেও এর উপস্থিতি ছিল সাধারণ ঘটনা। বর্তমানে প্রাপ্ত জীবাশ্ম নমুনাগুলোর উপর নির্ভর করে অনুমান করা হয় পালকের উপস্থিতি ছিল সম্ভবত ডাইনোসরদের একটি উৎপত্তিগত সাধারণ বৈশিষ্ট্য, কিন্তু পরবর্তীকালে তা কোনো কোনো প্রজাতির দেহ থেকে লোপ পায়[১২০]। পালক বা পালকসদৃশ অঙ্গের প্রত্যক্ষ নিদর্শন পায়া গেছে বহুসংখ্যক জীবাশ্ম নমুনায়, যাদের মধ্যে সরিস্কিয়া এবং অর্নিথিস্কিয়া দুই গোষ্ঠীই পড়ে। প্রাথমিক নিঅর্নিথিস্কিয়ান হেটারোডন্টোসরিড[১২১], থেরোপড[১২২] এবং প্রাথমিক সেরাটোপ্সিয়ানদের দেহে সরল, শাখাযুক্ত পালকসদৃশ অঙ্গ ছিল। আধুনিক পাখির উড্ডয়ন সহায়ক পালকের মতো শিরাযুক্ত প্রকৃত পালকের সন্ধান পাওয়া গেছে শুধুমাত্র থেরোপডদের উপবিভাগ ম্যানির্যাপ্টোরাদের মধ্যে, যে উপবিভাগের অন্তর্গত ছিল ওভির্যাপ্টোরোসর, ট্রুডন্টিড, ড্রোমিওসরিড এবং পাখিরা[৩৯][১২৩]। 'পিকনোফাইবার' নামক বিশেষ এক প্রকার পালকসদৃশ অঙ্গের খোঁজ পাওয়া গেছে টেরোসরদের মধ্যেও[১২৪]। এ'থেকে ধারণা করা হয় পালক ছিল আভেমেটাটারসালিয়া জীবশাখার বৈশিষ্ট্য যা ডাইনোসরদের থেকেও প্রাচীন[১২০]।
আর্কিওপ্টেরিক্স-এর জীবাশ্ম প্রথমবার ডাইনোসর ও পাখিদের যোগসূত্র প্রকাশ্যে নিয়ে আসে। এটিকে অন্তর্বর্তী জীবাশ্ম বলে গণ্য করা হয়, অর্থাৎ উভয় প্রকার প্রাণীর (ডাইনোসর এবং পাখি) বৈশিষ্ট্যই এর দেহে দেখা যায়। ডারউইনের লেখা অরিজিন অব স্পিশিজ বইয়ের প্রকাশনার মাত্র দু'বছর পরেকার এই আবিষ্কার বিবর্তনীয় জীববিজ্ঞান এবং সৃষ্টিবাদীদের মধ্যে জায়মান বিতর্ককে প্রবলভাবে উসকে দেয়। এই আদিম পাখিটি এত বেশি ডাইনোসরের মতো দেখতে যে এর বার্লিন নমুনাটির মতো চারপাশে পালকের ছাপ না থাকার কারণে অপর একটি নমুনাকে কম্পসোগ্ন্যাথাস মনে করা হয়েছিল[১২৫]। ১৯৯০ এর দশক থেকে আরও অনেক পালকযুক্ত ডাইনোসরের অবশেষ উদ্ধার করা হয়েছে, যা তাদের সাথে পাখিদের সংযোগের প্রমাণকে আরও দৃঢ় করেছে। এই নমুনাগুলোর অধিকাংশই পাওয়া গেছে উত্তর-পূর্ব চীনের লিয়াওনিং অঞ্চলের 'য়িক্সিয়ান প্রস্তরক্ষেত্র' থেকে, যা ক্রিটেশিয়াসে একটি বিচ্ছিন্ন দ্বীপ মহাদেশের আকারে ছিল। পালকের সন্ধান এখনও পর্যন্ত বিচ্ছিন্ন কিছু অঞ্চলে পাওয়া গেলেও পৃথিবীর অন্যত্রও উড়তে অক্ষম ডাইনোসরেরা পালকযুক্ত হয়ে থাকার সম্ভাবনা আছে। জীবাশ্ম নমুনায় পালকের বিরলতার একটা কারণ হতে পারে এই যে পালক, চামড়া ইত্যাদি নরম কলাকোশ প্রকৃতির প্রতিকূলতা সহ্য করে জীবাশ্মীভবন পর্যন্ত টিকে থাকতে পারে না। তাই জীবাশ্ম হওয়ার আগেই অধিকাংশ নমুনা থেকে পালকের চিহ্ন মুছে যায়[১২৬]।
পালকযুক্ত ডাইনোসরদের আবিষ্কার ও সেই সংক্রান্ত বিবৃতির প্রক্রিয়াটি আদৌ মসৃণ ছিল না। এর সবচেয়ে তীব্র বিরোধী সম্ভবত দুই বিজ্ঞানী অ্যালান ফেদুচ্চিয়া এবং থিয়াগার্টেন লিংহ্যাম-সলিয়ার, যাঁদের বক্তব্য হল ডাইনোসর নমুনায় প্রাপ্ত তথাকথিত 'পালকের' চিহ্ন আসলে তাদের চামড়ার নিচে অবস্থিত কোলাজেন তন্তুর পচনের ফলে সৃষ্টি হয়েছে[১২৭][১২৮][১২৯]। অবশ্য এই মতবাদ অন্যান্য বিজ্ঞানীরা সমর্থন করেন না, এবং ফেদুচ্চিয়ার গবেষণার বৈজ্ঞানিকতা নিয়েও প্রশ্ন উঠেছে[১৩০]।
কঙ্কাল
সম্পাদনাপালক সাধারণভাবে পাখির বৈশিষ্ট্য বলে স্বীকৃত, তাই পালকযুক্ত ডাইনোসরদের সাধারণত ডাইনোসর ও পাখির অন্তর্বর্তী নমুনা হিসেবে দেখা হয়। কিন্তু পালকের পাশাপাশি তাদের কঙ্কালতন্ত্রের অনেকগুলো সাদৃশ্যও উভয় শ্রেণীর যোগসূত্রের জোরদার প্রমাণ। কঙ্কালের যে সমস্ত জায়গায় পাখি আর ডাইনোসরের সাদৃশ্য চোখে পড়ে সেগুলো হল ঘাড়, শ্রোণীচক্র, কবজি, পুরোবাহু ও ঊরশ্চক্র, ফারকুলা এবং ঊরঃফলক। ক্ল্যাডিস্টিক বিশ্লেষণের দ্বারা উভয়ের কঙ্কালের তুলনা করলে মিলগুলো আরও ভালো বোঝা যায়[১৩১]।
কোমল কলাতন্ত্র
সম্পাদনা
ওহাইও বিশ্ববিদ্যালয়ের গবেষক প্যাট্রিক ও. কনর-এর একটি গবেষণা অনুযায়ী বৃহদাকার মাংসাশী ডাইনোসরদের দেহে আধুনিক পাখিদের মতোই জটিল বায়ুথলির বিন্যাস ছিল। পাখিদের মতোই থেরোপড ডাইনোসরদের ফুসফুসও সম্ভবত তাদের হাড়ের মধ্যে অবস্থিত ফাঁপা প্রকোষ্ঠে বায়ু সঞ্চালন করত। ও. কনর বলেছেন, "যে বৈশিষ্ট্যটাকে আনুষ্ঠানিকভাবে পাখিদের একচেটিয়া বলে মনে করা হত তা আসলে অন্য আকারে পাখিদের পূর্বপুরুষদের মধ্যেও বিদ্যমান ছিল"[১৩২]। অনলাইন পত্রিকা প্লোস ওয়ানে ২০০৮ খ্রিঃ প্রকাশিত একটি নিবন্ধে বিজ্ঞানীরা এরোস্টিয়ন রিওকলোরাডেনসিস-এর বিবরণ দেন, যাদের কঙ্কাল ডাইনোসরের দেহে পাখির অনুরূপ শ্বাসতন্ত্রের সেরা প্রমাণ দেখায়। এরোস্টিয়নের কঙ্কালের সিটি স্ক্যান করে তার দেহগহ্বরে বায়ুথলির অবস্থানের প্রমাণ মিলেছে[১৩৩][১৩৪]।
আচরণগত প্রমাণ
সম্পাদনাঅন্যতম ট্রুডন্টিড মেই এবং সিনর্নিথয়েড-দের জীবাশ্ম প্রমাণ করে যে কোনো কোনো ডাইনোসর দুই হাতের ফঁকে মাথা গুঁজে ঘুমাত[১৩৫]। মাথার উষ্ণতা সংরক্ষণ করার এই পদ্ধতি বর্তমানে পাখিদের মধ্যে দেখা যায়। অনেক রকম ডাইনোনিকোসর ও ওভির্যাপ্টোরোসর-এর নমুনাকে পাখির মতো বাসার উপর ডিমে তা দেওয়ার ভঙ্গীতে বসে থাকতে দেখা গেছে[১৩৬]। ডিমের আয়তন ও পূর্ণবয়স্ক ডাইনোসরদের দেহের ভরের অনুপাত বিশ্লেষণ করে বোঝা যায় এই সমস্ত ডিমে অধিকাংশ ক্ষেত্রে তা দিত পুরুষেরা, আর আধুনিক উড়তে অক্ষম পাখিদের মতোই তাদের বাচ্চারা স্বাবলম্বী হয়ে উঠত খুব তাড়াতাড়ি[১৩৭]। কোনো কোনো ডাইনোসর আধুনিক পাখিদের মতো গিজার্ডে পাথর রাখত। এই পাথরগুলো পৌষ্টিকতন্ত্রে সাহায্য করতে তথা পাকস্থলিতে শক্ত খাবার ও তন্তু প্রবেশ করলে তা থেঁতলে পরিপাকের উপযোগী করতে প্রয়োজন হত। জীবাশ্মে পাওয়া গিজার্ডের পাথরকে গ্যাস্ট্রোলিথ বলা হয়[১৩৮]।
প্রধান গোষ্ঠীসমূহের বিলুপ্তি
সম্পাদনাপাখিরা যে এক প্রকার ডাইনোসর এই আবিষ্কারের ফলে বোঝা গেছে জনপ্রিয় ধারণা অনুযায়ী ডাইনোসরেরা আসলে সম্পূর্ণ বিলুপ্ত হয়ে যায়নি[১৩৯]। অবশ্য সমস্ত উড়তে অক্ষম ডাইনোসর এবং অনেক প্রজাতির পাখি বাস্তবিকই আজ থেকে ৬ কোটি ৬০ লক্ষ বছর আগে বিলুপ্ত হয়ে যায়। অ্যামোনাইট, মোসাসর, প্লিসিওসর, টেরোসর ও বেশ কিছু স্তন্যপায়ী প্রভৃতি নানা গোত্রের অন্যান্য প্রাণীরাও ঐ সময়ে বিলুপ্ত হয়ে গিয়েছিল[৫]। বিশেষভাবে উল্লেখ্য ঘটনা হল এই যে সন্ধিপদীরা এই ঘটনার দ্বারা খুব একটা ক্ষতিগ্রস্ত হয়নি, ফলে তাদের ভক্ষণ করে অবশিষ্ট জীবিত প্রাণীকুলের খাদ্যের সরবরাহ অটুট থেকেছিল। এই মহাবিলুপ্তির ঘটনা ক্রিটেশিয়াস-প্যালিওজিন বিলুপ্তি ঘটনা নামে পরিচিত। ১৯৭০ এর দশক থেকে এই ঘটনার প্রকৃতি নিয়ে প্রচুর গবেষণা ও আলোচনা হয়েছে। বর্তমানে একাধিক তত্ত্বের একটি সম্মিলিত প্রভাবের কথা পুরাজীববিদরা সমর্থন করেন। একটি উল্কার সাথে পৃথিবীর সংঘর্ষের ঘটনাকে প্রাথমিক বা প্রত্যক্ষ কারণ হিসেবে মানা হলেও কোনো কোনো বিজ্ঞানী ডাইনসরদের জীবাশ্ম প্রমাণ বিলোপের অন্যান্য সম্ভাবনার কথা তুলে ধরেছেন[১৪০][১৪১][১৪২]।
মেসোজোয়িক মহাযুগে বায়ুমণ্ডলের গঠন সম্বন্ধে বেশ কিছু গুরুত্বপূর্ণ তত্ত্ব আছে যা এই বিলুপ্তি ঘটনার উপর আলোকপাত করতে পারে। কেউ কেউ বলেন বাতাসে অক্সিজেনের মাত্রা তখন বর্তমানের তুলনায় অনেক বেশি ছিল; অন্যেরা বলেন ডাইনোসর ও পাখিদের বিভিন্ন প্রজাতির শ্বাসতন্ত্র যেমন উন্নত অভিযোজনের সাক্ষ্য বহন করে তা বাতাসে অক্সিজেন বেশি থাকলে প্রয়োজনই হত না[১৪৩]। ক্রিটেশিয়াসের অন্তিম লগ্নে পরিবেশে ব্যাপক পরিবর্তন ঘটছিল। বিভিন্ন আগ্নেয়গিরির সক্রিয়তা হ্রাস পাচ্ছিল, ফলে বায়ুমণ্ডলে কার্বন ডাই অক্সাইডের মাত্রা কমে গিয়ে জলবায়ু শীতল হতে আরম্ভ করে। অক্সিজেনের পরিমাণও কমে যেতে থাকে। অনেক বিজ্ঞানী ধারণা করেন জলবায়ুর পরিবর্তন ও অক্সিজেনের স্বল্পতা মিলিতভাবে বহু প্রজাতির বিলুপ্তির জন্য দায়ী ছিল[১৪৪]।
মহাজাগতিক সংঘর্ষ তত্ত্ব
সম্পাদনা
১৯৮০ খ্রিঃ ওয়াল্টার আলভারেজ এবং তার সহকর্মীরা গ্রহাণু সংঘর্ষ তত্ত্বের অবতারণা করেন। এই তত্ত্বে ডাইনোসরদের অবলুপ্তিকে পৃথিবীর সাথে একটি অত্যুজ্জ্বল উল্কা বা বোলাইডের সংঘর্ষের ফল হিসেবে তুলে ধরা হয়। ঘটনাটা ঘটেছিল কমবেশি ৬ কোটি ৬০ লক্ষ বছর আগে। আলভারেজ ও তার সহকর্মীরা বলেন যে বিশ্বের নানা প্রান্ত থেকে প্রাপ্ত সমসাময়িক পাথরের স্তরে অতিরিক্ত মাত্রায় ইরিডিয়ামের উপস্থিতি এই সংঘর্ষের প্রত্যক্ষ প্রমাণ[১৪৫]। এখন পর্যন্ত প্রাপ্ত নিদর্শন থেকে বোঝা যায় ঐ সময় প্রায় ৫ থেকে ১৫ কিলোমিটার (৩ থেকে ৯ মাইল) ব্যাসের একটি উল্কা মেক্সিকোর দক্ষিণ-পূর্বে অবস্থিত য়ুকাটান উপদ্বীপে আছড়ে পড়ে বর্তমান ১৮০ কিমি (১১০ মা) চিক্সুলাব উল্কাখাত গঠনপূর্বক মহাবিলুপ্তি ত্বরান্বিত করে[১৪৬][১৪৭]। ডাইনোসরেরা এই সংঘর্ষের অব্যবহিত আগে বর্ধিষ্ণু ছিল না ক্ষয়িষ্ণু ছিল তা নিয়ে বিজ্ঞানীরা নিশ্চিত নন। কেউ কেউ বলেন সংঘর্ষের পর পৃথিবীব্যাপী বায়ুমণ্ডলের তাপমাত্রা অস্বাভাবিকভাবে হ্রাস পেয়েছিল আর এই অবস্থা দীর্ঘ সময় ধরে বজায় ছিল। অপরেরা বলেন বরং সংঘর্ষের সাথে সাথে পৃথিবীব্যাপী অস্বাভাবিক তাপপ্রবাহের সৃষ্টি হওয়ার সম্ভাবনাই বেশি। যে সমস্ত বিজ্ঞানী সংঘর্ষ তত্ত্বের সমর্থক তারা এখন মোটামুটি একটা ঐকমত্য্যে এসেছেন এই বলে যে উল্কাটি প্রত্যক্ষ (সংঘর্ষের ফলে উৎপন্ন তাপের মাধ্যমে) ও পরোক্ষ (সংঘর্ষের ফলে ভূ-ত্বক থেকে ছিটকে ওঠা পদার্থকণা সূর্যরশ্মির অধিকাংশ মহাকাশে প্রতিফলিত করে দিয়ে পৃথিবীকে ঠান্ডা করে দেওয়ার ফলে) - দুই ভাবেই মহাবিলুপ্তির পথ সুগম করেছিল। যদিও শুধু জীবাশ্ম প্রমাণের উপর নির্ভর করে কত তাড়াতাড়ি এই বিলুপ্তি হয়েছিল তা বলা যায় না, তবুও সামগ্রিক বিবেচনাপ্রসূত একাধিক মডেল অনুযায়ী গোটা ঘটনাটা ঘটতে কয়েক বছর নয়, বরং কয়েক ঘণ্টা মাত্র লেগেছিল বলে অনুমান করা হয়[১৪৮]।
ডেকান ট্র্যাপ
সম্পাদনা২০০০ খ্রিষ্টাব্দের আগে পর্যন্ত যে সমস্ত মতবাদ ডেকান ট্র্যাপের ব্যাসল্ট প্লাবনের ঘটনাটিকে মহাবিলুপ্তির সাথে সংযুক্ত হিসেবে ব্যাখ্যা করত সেগুলো একটা ব্যাপারে একমত ছিল, আর তা হল এই যে এই বিলুপ্তি ঘটেছিল ধীরে ধীরে। এর কারণ দাক্ষিণাত্যে ব্যাসল্ট প্লাবন শুরু হয় আজ থেকে ৬ কোটি ৮০ লক্ষ বছর আগে এবং তা চলতে থাকে ২০ লক্ষ বছর ধরে। কিন্তু এই প্রমাণও আছে যে সমগ্র দাক্ষিণাত্য লাভা মালভূমির দুই তৃতীয়াংশই তৈরি হয়েছিল মহাবিলুপ্তির আগের ১০ লক্ষ বছরে, কাজেই এই বিস্ফোরণগুলোর সময় দ্রুত অনেক প্রজাতির বিলোপনের সম্ভাবনা আছে; কয়েক হাজার বছরের মধ্যেই তা হয়ে থাকতে পারে। কিন্তু একক সংঘর্ষের ফলে ঘটিত মহাবিলুপ্তির পক্ষে কয়েক হাজার বছরও একটু বেশিই সময়[১৪৯][১৫০]।
দাক্ষিণাত্য লাভা মালভূমি বা ডেকান ট্র্যাপ একাধিক পদ্ধতিতে মহাবিলুপ্তিতে সাহায্য করে থাকতে পারে। যেমন, এই অঞ্চলের ভূ-আলোড়নের ফলে বাতাসে ছাই, ধুলো আর সালফারঘটিত এরোসল যোগ হওয়ার মাধ্যমে সূর্যরশ্মির প্রবেশপথ আটকে গিয়ে উদ্ভিদকুলের সালোকসংশ্লেষ সাময়িকভাবে বন্ধ হয়ে গিয়ে থাকতে পারে। এছাড়া দাক্ষিণাত্য মালভূমির লাভা নিঃসরণের সাথে সাথে প্রচুর পরিমাণ কার্বন ডাই অক্সাইড গ্যাস নির্গত হয়ে থাকতে পারে, যা ছিটকে ওঠা ধুলো ইত্যাদি থিতিয়ে যাওয়ার পর অত্যধিক মাত্রায় বায়ুমণ্ডলের গ্রিনহাউস প্রভাব বাড়িয়ে পৃথিবীকে হয়তো গরম করে তুলেছিল[১৫০]। বাস্তবিক, ডাইনোসরদের বিলুপ্তির আগে দাক্ষিণাত্য লাভা মালভূমি গঠনের সময় নির্গত গ্যাসের ফলে পৃথিবীব্যাপী প্রবল উষ্ণায়ন হয়েছিল। কোনো কোনো সূত্র অনুযায়ী "চিক্সুলাব উল্কাখাত গঠনের পাঁচ লক্ষ বছর আগেই বায়ুমণ্ডলের গড় উষ্ণতা ৮ °C (১৪ °F) বৃদ্ধি পেয়েছিল"[১৪৯][১৫০]।
যখন বিজ্ঞানীদের মধ্যে ডেকান ট্র্যাপ ঘটিত ধীর মহাবিলুপ্তির তত্ত্ব ক্রমশ গ্রহণযোগ্যতা পাচ্ছিল, তখন লুই ওয়াল্টার আলভারেজ বলেন এই তত্ত্ব কিছু বিচ্ছিন্ন সূত্রকে অত্যধিক গুরুত্ব আরোপ করে গবেষণা ভুল পথে নিয়ে যাচ্ছে। প্রথম দিকে তার মতামতকে পাত্তা দেওয়া না হলেও পরবর্তী গবেষণায় তার তত্ত্বের গ্রহণযোগ্যতা প্রমাণিত হয়েছে। অর্থাৎ, ডাইনোসরদের বিলুপ্তি যে আকস্মিক কোনো ঘটনা (সংঘর্ষ তত্ত্ব) সেই মতবাদই এখন গ্রাহ্য বলা যায়। তবে আলভারেজ স্বয়ং স্বীকার করেছেন যে সংঘর্ষের আগে পৃথিবীতে আরও কয়েকটি আমূল পরিবর্তন সাধিত হয়েছিল, যথা সমুদ্রতলের উচ্চতা হ্রাস এবং ক্রমিক অগ্ন্যুৎপাতের মাধ্যমে দাক্ষিণাত্য মালভূমির সৃষ্টি; এবং এই ঘটনাগুলোও মহাবিলুপ্তির অনুঘটক হয়ে থাকতে পারে[১৫১]।
প্যালিওসিনে সম্ভাব্য ধারাবাহিকতা
সম্পাদনাকখনও কখনও ক্রিটেশিয়াস-প্যালিওজিন সীমার উপর থেকে উড়তে অক্ষম ডাইনোসরদের অবশেষ পাওয়া গেছে। ২০০১ খ্রিঃ পুরাজীববিদ জিয়েলেন্স্কি এবং বুদান নিউ মেক্সিকোর সান জুয়ান উপত্যকা থেকে প্রাপ্ত একটি হ্যাড্রোসরের একক পায়ের হাড়ের বর্ণনা দেন এবং এটিকে প্যালিওসিন ডাইনোসরদের অস্তিত্বের প্রমাণ হিসেবে তুলে ধরেন। যে প্রস্তরক্ষেত্র থেকে হাড়টি পাওয়া গিয়েছিল তার জন্ম প্যালিওসিন উপযুগে, আনুমানিক ৬ কোটি ৪৫ লক্ষ বছর আগে। যদি ঐ হাড়টি আবহবিকারের ফলে ঐ পাথরের স্তরে এসে না আটকে গিয়ে থাকে, তাহলে ওটি প্রকৃতই ক্রিটেশিয়াস-প্যালিওজিন বিলুপ্তি ঘটনার প্রায় ৫ লক্ষ বছর পরে সিনোজোয়িক মহাযুগেও পৃথিবীতে ডাইনোসরদের অস্তিত্বের পক্ষে প্রমাণ দেয়[১৫২]। অন্যান্য প্রমাণের অন্তর্গত হল হেল ক্রিক প্রস্তরক্ষেত্র থেকে প্রাপ্ত ডাইনোসর অবশেষ যা ক্রিটেশিয়াস-প্যালিওজিন সীমার ১.৩ মিটার (৫১ ইঞ্চি) উপরে (৪০০০০ বছর পরে) অবস্থিত। চীন সমেত পৃথিবীর অন্যান্য জায়গা থেকেও অনুরূপ প্রমাণ মিলেছে[১৫৩]। অনেক বিজ্ঞানী অবশ্য এই সমস্ত প্রমাণকে জালি বলে অস্বীকার করেছেন, তাদের মতে এগুলো পূর্ববর্তী পাথরের স্তর থেকে আবহবিকারের ফলে স্থানচ্যুত হয়ে পরবর্তী যুগের স্তরে আটকা পড়েছে[১৫৪][১৫৫]। যদিও হাড়গুলোর বয়স নির্ণয়ের প্রত্যক্ষ পদ্ধতিতে পরবর্তী সময়কালেরই সমর্থন পাওয়া যায়; ইউরেনিয়াম-দস্তা (U-Pb) তারিখ গণনাপদ্ধতিতে এগুলোর বয়স নির্ণীত হয়েছে ঠিক ৬ কোটি ৪৮ লক্ষ ± ৯০ লক্ষ বছর[১৫৬]। সঠিক হলেও অবশ্য এই সমস্ত প্রমাণ মহাবিলুপ্তির ঘটনার প্রাবল্যের খুব একটা তারতম্য প্রতিষ্ঠা করতে পারবে না[১৫৪]।
চর্চার ইতিহাস
সম্পাদনাডাইনোসর জীবাশ্মের সঙ্গে মানুষের পরিচয় হয়েছিল হাজার হাজার বছর আগেই, যদিও তারা জিনিসগুলোর প্রকৃতি সম্বন্ধে অবগত ছিল না। চীন দেশে ডাইনোসরের আধুনিক প্রতিশব্দ কংলং ( 恐龍, বা 'ভয়ানক ড্রাগন'), আর ডাইনোসরের হাড়কেও চীনারা ড্রাগনের হাড় বলেই মনে করতেন। যেমন, পশ্চিম জিন্ রাজবংশের শাসনকালে (২৬৫-৩১৬ খ্রিঃ) চাং কু-এর লেখা বই হুয়া য়াং গুও ঝি-তে সিচুয়ান প্রদেশের উচেনে ড্রাগনের হাড় আবিষ্কারের কথা আছে[১৫৭]। মধ্য চীনের গ্রামবাসীরা অনেককাল ধরেই ওষুধ প্রস্তুত করতে 'ড্রাগনের হাড়' ব্যবহার করেন এবং এই ঐতিহ্য আজও বজায় আছে[১৫৮]। ইউরোপে ডাইনোসরের হাড়কে পৌরাণিক দৈত্য বা বাইবেলে বর্ণিত অলৌকিক জীবসমূহের হাড় বলে ধারণা করা হত[১৫৯]।

ডাইনোসরের হাড়ের আধুনিক বৈজ্ঞানিক পর্যালোচনা ও বিবরণ দেওয়ার প্রক্রিয়া প্রথম শুরু হয় সপ্তদশ শতাব্দীর ইংল্যান্ডে। ১৬৭৬ খ্রিঃ অক্সফোর্ডশায়ারের চিপিং নর্টনের নিকটবর্তী কর্নওয়েলের কাছে একটি চুনাপাথরের স্তরে একটি হাড় পাওয়া যায়, যেটি পরবর্তীকালে মেগালোসরাসের একটি উরুর হাড় বলে প্রমাণিত হয়েছে[১৬০]। হাড়টি পাওয়ার পর অক্সফোর্ড বিশ্ববিদ্যালয়ের রসায়নের অধ্যাপক এবং আশমোলিয়ান মিউজিয়ামের প্রথম কিউরেটর রবার্ট প্লটের কাছে পাঠানো হয়। তিনি ১৬৭৭ খ্রিঃ প্রকাশিত তার বই ন্যাচারাল হিস্ট্রি অফ অক্সফোর্ডশায়ার-এ এটির বিস্তারিত বর্ণনা দেন। তিনি সঠিকভাবে অনুমান করেন যে হাড়টি প্রকৃতপক্ষে কোনো বড় আয়তনের প্রাণীর উরুর হাড়ের নিম্নাংশ, এবং এ-ও বলেন যে বর্তমানে জ্ঞাত কোনো প্রজাতির পক্ষে এটির আয়তন বড্ড বেশি। তাই তিনি সিদ্ধান্ত করেন যে এটি আসলে বাইবেলে বর্ণিত দৈত্যাকার মানুষদের কারো পায়ের হাড়। ১৬৯৯ খ্রিঃ স্যার আইজাক নিউটনের বন্ধু এডওয়ার্ড লুইড, অক্সফোর্ডশায়ারের উইটনির নিকটে ক্যাসওয়েলে প্রাপ্ত[১৬১] একটি সরোপডের দাঁতের "রুটেলাম ইমপ্লিকেটাম" নাম দিয়ে যে নিবন্ধটি প্রকাশ করেন সেটিকে ডাইনোসর সংক্রান্ত প্রথম বৈজ্ঞানিক দৃষ্টিভঙ্গী থেকে লেখা নিবন্ধ বলা যেতে পারে[১৬২][১৬৩]।

১৮১৫ থেকে ১৮২৪ এর মধ্যে অক্সফোর্ড বিশ্ববিদ্যালয়ের ভূতত্ত্বের অধ্যাপক রেভারেন্ড উইলিয়াম বাকল্যান্ড মেগালোসরাসের আরও হাড় সংগ্রহ করে কোনো বৈজ্ঞানিক পত্রিকায় প্রথম কোনো ডাইনোসরের বিবরণ দেওয়ার কৃতিত্ব অর্জন করেন[১৬০][১৬৪]। এর পরে প্রাপ্ত ডাইনোসর ছিল ইগুয়ানোডন। ১৮২২ খ্রিঃ এর অবশেষ খুঁজে পেয়েছিলেন ম্যারি অ্যান ম্যান্টেল, ইংরেজ ভূতত্ত্ববিদ গিডিয়ন ম্যান্টেলের স্ত্রী। গিডিয়ন ম্যান্টেল এই জীবাশ্ম এবং আধুনিক ইগুয়ানার হাড়ের মধ্যে সাদৃশ্য খুঁজে পান। তার অনুসন্ধানের ফল ১৮২৫ খ্রিঃ প্রকাশিত হয়[১৬৫][১৬৬]।
এই সমস্ত "বৃহৎ সরীসৃপ জীবাশ্ম" সংক্রান্ত গবেষণা শীঘ্রই ইউরোপীয় ও আমেরিকান বিজ্ঞানীদের কাছে চিত্তাকর্ষক হয়ে দাঁড়ায়, আর ১৮৪২ খ্রিঃ ইংরেজ পুরাজীববিদ রিচার্ড ওয়েন 'ডাইনোসর কথাটির প্রচলন করেন। তিনি লক্ষ্য করেন যে তখনও পর্যন্ত উদ্ধার করা তিনটি প্রজাতির জীবাশ্ম, যথা ইগুয়ানোডন, মেগালোসরাস ও হাইলিয়সরাস-এর একাধিক সাধারণ বৈশিষ্ট্য রয়েছে এবং সেই সূত্রে তাদের একটি নির্দিষ্ট শ্রেণিবিন্যাসগত বিভাগের আওতায় আনার ব্যবস্থা করেন। রাণী ভিক্টোরিয়ার স্বামী রাজকুমার অ্যালবার্টের পৃষ্ঠপোষকতায় ওয়েন লন্ডনের দক্ষিণ কেনসিংটনে ন্যাচারাল হিস্ট্রি মিউজিয়াম প্রতিষ্ঠা করেন। প্রাথমিকভাবে এই জাদুঘর নির্মাণের উদ্দেশ্য ছিল ডাইনোসর জীবাশ্ম ও অন্যান্য জীববিদ্যা ও ভূতত্ত্ব সংক্রান্ত দ্রষ্টব্যের প্রদর্শনের ব্যবস্থা করা[১৬৭]।
১৮৫৮ খ্রিঃ বিজ্ঞানী ফুল্কের তত্ত্বাবধানে নিউ জার্সির হ্যাডনফীল্ড শহরের মার্ল থেকে প্রথম আমেরিকান ডাইনোসর আবিষ্কৃত হয়। সে'দেশের অন্যত্র তার আগে ডাইনোসর জীবাশ্ম পাওয়া গেলেও সেগুলোর সম্বন্ধে সঠিক ধারণা গড়ে ওঠেনি। এই প্রজাতিটির নাম দেওয়া হয় হ্যাড্রোসরাস ফাউলকিয়াই। এই আবিষ্কারটির গুরুত্ব অনেক, কারণ এটি ডাইনোসরদের প্রাপ্ত প্রথম সম্পূর্ণ কঙ্কালগুলোর মধ্যে অন্যতম,আর এটা থেকে পরিষ্কার বোঝা গিয়েছিল যে জীবিত অবস্থায় প্রাণীটি ছিল দ্বিপদ। এর আগে পর্যন্ত বিজ্ঞানীরা মনে করতেন ডাইনোসরেরা অন্যান্য সরীসৃপদের মতোই চার পায়ে হাঁটত। ফুল্কের আবিষ্কার মার্কিন যুক্তরাষ্ট্রে ডাইনোসর নিয়ে উন্মাদনার সৃষ্টি করে[১৬৮]।


ডাইনোসর উন্মাদনার চূড়ান্ত উদাহরণ হল দুই বিজ্ঞানী এডওয়ার্ড ড্রিঙ্কার কোপ এবং অথনিয়েল চার্লস মার্শের তিন দশক ব্যাপী গবেষণার লড়াই, যা বর্তমানে জীবাশ্ম যুদ্ধ বা "অস্থিযুদ্ধ" নামে খ্যাত। এঁরা দু'জনেই সবচেয়ে বেশি সংখ্যায় নতুন ডাইনোসর আবিষ্কারের কৃতিত্বের দাবীদার হতে চাইতেন। এঁদের প্রতিদ্বন্দ্বিতার সূত্রপাত হয় খুব সম্ভবত মার্শের লেখা একটি নিবন্ধের মাধ্যমে, যেখানে মার্শ দেখান যে কোপ একটি ইলাসমোসরাস-এর কঙ্কালের ভুল পুনর্নির্মাণ করেছেন; কোপ ঐ প্লিসিওসরটির মাথা তার লেজের জায়গায় বসিয়েছিলেন। ত্রিশ বছর ধরে চলার পর ১৮৯৭ খ্রিঃ কোপের মৃত্যুর মাধ্যমে জীবাশ্ম যুদ্ধ শেষ হয়। মৃত্যুকালে কোপ তার সঞ্চয়ের সম্পূর্ণ অংশ ডাইনোসর শিকারে নিয়োজিত করে দিয়েছিলেন। মার্শ এই যুদ্ধ 'জেতেন' মূলত মার্কিন ভূতাত্ত্বিক সর্বেক্ষণের সাথে তার একটি চুক্তি হয়ে থাকার কারণে; এর ফলে তার গবেষণা ও অনুসন্ধানে অর্থের বরাদ্দ নিয়মিত ছিল। দুঃখের বিষয় এই যে, এঁদের দু'জনের অনুঃসৃত পদ্ধতির ত্রুটির জন্য অনেক ডাইনোসর নমুনা ক্ষতিগ্রস্ত হয়েছিল; যেমন, তাদের দল অনেক সময় পাহাড় থেকে নমুনা উদ্ধার করতে ডায়নামাইট ব্যবহার করত (আধুনিক কালে পুরাজীববিদরা ভুলেও এ'কাজ করেন না)। অবশ্য পদ্ধতিগত ত্রুটি সত্ত্বেও পুরাজীববিদ্যায় কোপ ও মার্শের অবদান বিপুল; মার্শ ডাইনোসরদের ৮৬টি নতুন প্রজাতি আবিষ্কার করেন এবং কোপ আবিষ্কার করেন ৫৬টি, অর্থাৎ সর্বমোট ১৪২ টি নতুন প্রজাতি এই দুই বিজ্ঞানীর চেষ্টায় আবিষ্কৃত হয়। কোপের সংগ্রহ বর্তমানে নিউ ইয়র্কের আমেরিকান মিউজিয়াম অফ ন্যাচারাল হিস্ট্রি-তে সংরক্ষিত আছে, আর মার্শের আবিষ্কারগুলো রক্ষিত আছে ইয়েল বিশ্ববিদ্যালয়ের পীবডি মিউজিয়াম অফ ন্যাচারাল হিস্ট্রি-তে[১৬৯]।
১৮৯৭ খ্রিঃ এর পর অ্যান্টার্কটিকা সমেত সারা পৃথিবী জুড়ে ডাইনোসরের জীবাশ্মের খোঁজ শুরু হয়। প্রথম আবিষ্কৃত দক্ষিণ মেরু ডাইনোসর হল অন্যতম অ্যাঙ্কিলোসরিড অ্যান্টার্কটোপেল্টা অলিভেরোই। এর জীবাশ্ম পাওয়া গিয়েছিল ১৯৮৬ খ্রিঃ জেম্স রস দ্বীপ থেকে[১৭০], যদিও প্রথম দক্ষিণ মেরু মহাদেশীয় ডাইনোসর প্রজাতি হিসেবে কোনো বৈজ্ঞানিক পত্রিকায় প্রথম বর্ণনা দেওয়া হয় থেরোপড ক্রায়োলোফোসরাস এলিয়টির, ১৯৯৪ খ্রিঃ[১৭১]।
বর্তমানে জ্ঞাত ডাইনোসর জীবাশ্মের 'হটস্পট', অর্থাৎ অত্যধিক সংখ্যায় সংরক্ষিত জীবাশ্ম পাওয়া যায় এমন অঞ্চল, হল দক্ষিণ আমেরিকার দক্ষিণাঞ্চল (বিশেষত আর্জেন্টিনা) এবং চীন। চীন দেশ থেকে অনেক সংখ্যক বিরল প্রজাতির পালকযুক্ত ডাইনোসর-এর সন্ধান পাওয়া গেছে; প্রধানত জীবাশ্মক্ষেত্রগুলোর সুবিধেজনক ভূমিরূপের কারণে আর প্রাচীনকালে এই অঞ্চলের শুষ্ক জলবায়ুর কারণে, যা জীবাশ্মীভবনের অনুকূল ছিল[১২৬]।
"ডাইনোসর নবজাগরণ"
সম্পাদনা
১৯৭০ এর দশক থেকে এখনও পর্যন্ত ডাইনোসর সংক্রান্ত গবেষণার জগতে উৎসাহ ও নৈপুণ্যের প্রাবল্য দেখা যাচ্ছে। এই পরিবর্তনের সূত্রপাত হয়েছিল জন অস্ট্রমের ডাইনোনিকাস আবিষ্কারের দ্বারা। এই ডাইনোসরটি যে জীবৎকালে অতিমাত্রায় সক্রিয় এবং খুব সম্ভবত উষ্ণশোণিত ছিল, তা বিজ্ঞানীরা এর আবিষ্কারের সময়ই অনুমান করেছিলেন। ফলে ডাইনোসরদের অনুষ্ণশোণিত মনে করার ধারণাটা বাতিল হয়ে যায়। পুরাজীববিদ্যার মেরুদন্ডী সংক্রান্ত শাখাটি এই আবিষ্কারের ফলে প্রবলভাবে আলোড়িত হয় এবং বর্তমানে এটি বিজ্ঞান হিসেবে আন্তর্জাতিক মহলের মনোযোগ আকর্ষণ করেছে। ভারত, দক্ষিণ আমেরিকা, মাদাগাস্কার, অ্যান্টার্কটিকা ও বিশেষ করে চীন প্রভৃতি ইতঃপূর্বে খনন না করা অনেক অঞ্চল থেকে গুরুত্বপূর্ণ বহু ডাইনোসর আবিষ্কৃত হয়েছে ও এখনও হয়ে চলেছে। চীনে প্রাপ্ত পালকযুক্ত ডাইনোসরেরা পাখি ও ডাইনোসরের বিবর্তনীয় যোগসূত্রের জোরালো প্রমাণ হিসেবে উঠে এসেছে। বিভিন্ন জীবগোষ্ঠীর পারস্পরিক সম্পর্ককে যা অত্যাধুনিক পদ্ধতি অবলম্বন করে বিচার করে, সেই ক্ল্যাডিস্টিক্স বিদ্যার বহুল প্রয়োগের ফলে ডাইনোসরেদের শ্রেণিবিন্যাসের সুবিধে হয়েছে অনেক। জীবাশ্ম প্রমাণ প্রায়শই অসম্পূর্ণ থেকে যায় বলে বিভিন্ন ডাইনোসর গোষ্ঠীর বিবর্তনের প্রবাহ নির্ণয়ের ক্ষেত্রে ক্ল্যাডিস্টিক্সের সাহায্য নেওয়া হয়[১৭২]।
কোমল কলাতন্ত্র ও ডিএনএ
সম্পাদনাজীবাশ্মীভূত ডাইনোসরের দেহে কোমল কলাতন্ত্রের অবস্থানের অন্যতম উৎকৃষ্ট নমুনা পাওয়া গেছে ইতালির পিয়েত্রারোজায়। ১৯৯৮ খ্রিঃ নিবন্ধীকৃত এই আবিষ্কারটি অন্যতম সিলুরোসর সিপিওনিক্স স্যাম্নিটিকাস প্রজাতির একটি ক্ষুদ্র ও অপ্রাপ্তবয়স্ক সদস্যের। নমুনাটিতে ক্ষুদ্রান্ত্র, কোলন, যকৃৎ, পেশি ও শ্বাসনালীর অংশবিশেষ সংরক্ষিত ছিল[৫১]।
সায়েন্স পত্রিকার ২০০৫ খ্রিঃ মার্চ মাসে প্রকাশিত সংখ্যায় পুরাজীববিদ মেরি হিগবি শোয়াইৎজার এবং তার দল একটি ৬ কোটি ৮০ লক্ষ বছর পুরোনো টির্যানোসরাস রেক্সের পায়ের হাড়ে নমনীয় পদার্থের চিহ্ন প্রাপ্তির কথা ঘোষণা করেন, যে পদার্থ প্রকৃত কোমল কলাতন্ত্রের অবশেষরূপে নির্দিষ্ট করা হয়েছে। হাড়টি পাওয়া গিয়েছিল মন্টানার হেল ক্রিক প্রস্তরক্ষেত্র থেকে, এবং উক্ত বিজ্ঞানীরা নমুনাটি পাওয়ার পর সিদ্ধান্তে উপনীত হওয়ার আগে তাতে জলসঞ্চার করেন[৫২]।
জীবাশ্মীভূত হাড়টির মজ্জাগহ্বর থেকে প্রথমে কয়েক সপ্তাহ ধরে খনিজ পদার্থ নিষ্কাশন করা হয়। এই পদ্ধতির নাম 'ডি-মিনারেলাইজেশন'। এর পর শোয়াইৎজার তার মধ্যে রক্তবাহ, অস্থি ধাত্র এবং যোগ কলা (অস্থি তন্তু) প্রভৃতির সুসংরক্ষিত অবশেষ দেখতে পান। অণুবীক্ষণ যন্ত্রের তলায় পুঙ্খানুপুঙ্খ অনুসন্ধানের পর দেখা গেছে আলোচ্য ডাইনোসর হাড়টি এমনকি কোশীয় স্তরেও বিভিন্ন সূক্ষ্ম গঠন অবিকৃত রেখে দিয়েছে! এই কোশীয় উপাদানগুলোর প্রকৃতি ও গঠনপদ্ধতি এখনও যথাযথভাবে নির্ণয় করা হয়নি; তাই শোয়াইৎজারের গবেষণার পূর্ণ অভিঘাত সম্পর্কে এখনও নিশ্চিত হওয়া যায়নি। এই গবেষণা এখনও জারি আছে[৫২]।
২০০৯ খ্রিঃ শোয়াইৎজার ও তার দল সতর্ক গবেষণার পর ঘোষণা করেন যে অন্যান্য বেশ কিছু নমুনায় তারা অনুরূপ অবস্থা লক্ষ করেছেন। এগুলোর মধ্যে ছিল মন্টানারই জুডিথ নদী প্রস্তরক্ষেত্র থেকে প্রাপ্ত একটি হ্যাড্রোসরিড ও একটি ব্র্যাকিলোফোসরাস ক্যানাডেনসিস। এই নমুনাগুলোয় আরও সূক্ষ্ম কলাকোশের সংরক্ষণের চিহ্ন রয়েছে; এমনকি নিউক্লিয়াসের চিহ্নযুক্ত অবিকৃত অস্থিকোশ ও লোহিত রক্তকণিকা পর্যন্ত। এছাড়া নমুনাগুলোয় পাওয়া গেছে কোলাজেন, যা প্রথম প্রাপ্ত টির্যানোসরাসটির দেহেও ছিল। প্রাণীদেহের কোলাজেনের প্রকৃতি তার ডিএনএ অনুযায়ী পরিবর্তিত হয় এবং এই সমস্ত ডাইনোসরের কোলাজেনের প্রকৃতি আধুনিক মুরগি ও উটপাখির কোলাজেনের অনুরূপ[১৭৩]।
জীবাশ্মীভূত ডাইনোসরের দেহ থেকে সফলভাবে প্রাচীন ডিএনএ নিষ্কাশনের প্রচেষ্টার খবর পাওয়া গেছে দু'বার, কিন্তু এগুলোর সত্যতা যাচাই করা হয়নি।[১৭৪] যদিও পাখি, সরীসৃপ প্রভৃতি আধুনিক প্রজাতিদের জিন বিন্যাসের সাথে তুলনা করে বিশ্লেষণাত্মক জাতিজনি পুনর্গঠনের সময় একটি আনুমানিক ডাইনোসরের দেহের একটি আনুমানিক সক্রিয় পেপটাইড সম্পর্কে ধারণা করা হয়েছে।[১৭৫] এছাড়া অনেক ডাইনোসর নমুনায় হিমোগ্লোবিন[১৭৬] সমেত অনেক প্রোটিনের সন্ধান পাওয়ার খবর এসেছে।[১৭৭][১৭৮]
মানুষের সংস্কৃতিতে উপস্থাপনা
সম্পাদনা

মানুষের নিরিখে অনেক ডাইনোসরই ছিল অদ্ভুত আকারবিশিষ্ট বিশাল আয়তনের প্রাণী। স্বাভাবিকভাবেই তারা আবিষ্কারের সময় থেকে জনমানসে প্রবল সাড়া ফেলে বর্তমানে সংস্কৃতির একটি অবিচ্ছেদ্য অঙ্গে পরিণত হয়েছে। 'ডাইনোসর' শব্দটি যে নিত্যদিনে ক্রমশ ব্যবহার্য হয়ে উঠছে, এই ঘটনাই প্রাণীগুলোর সাংস্কৃতিক গুরুত্ব নির্দেশ করে। সাধারণত যে সমস্ত জিনিস বা বিষয় খাপছাড়া রকম বিরাট, বর্তমানে যার প্রচলন আর নেই বা যেগুলো খুব তাড়াতাড়ি লোপাট হয়ে যাবে বলে ধরে নেওয়া হয় তাদের নির্দেশ করতে 'ডাইনোসর' কথাটা ব্যবহৃত হচ্ছে।[১৭৯]
ডাইনোসর সম্বন্ধে জনসাধারণের মনে উন্মাদনার প্রথম সঞ্চার ঘটে ভিক্টোরীয় ইংল্যান্ডে। ঐ যুগ চলাকালীন ১৮৫৪ খ্রিঃ অর্থাৎ ডাইনোসর জীবাশ্মসমূহের প্রথম বৈজ্ঞানিক বর্ণনার তিন দশক বাদে লন্ডনের ক্রিস্টাল প্যালেস পার্কের বিখ্যাত ডাইনোসর ভাস্কর্যগুলো জনসাধারণের দর্শনের জন্য উন্মোচিত হয়। ক্রিস্টাল প্যালেস ডাইনোসরেরা অল্প সময়ের মধ্যেই এত জনপ্রিয় হয়ে ওঠে যে ছোট ছোট অনুকৃতির চাহিদা অনুমান করে ডাইনোসর পুতুলের জমজমাট ব্যবসা শুরু হয়ে যায়। পরবর্তী দশকগুলোতে পৃথিবীর নানা জায়গায় জাদুঘরসমূহে ডাইনোসর সংক্রান্ত জমকালো বিভাগ প্রতিষ্ঠা করে তাদের অবশেষ ও অনুকৃতি ইত্যাদির প্রদর্শনীর ব্যবস্থা করা হয়; ফলে পরবর্তী প্রজন্মের সদস্যেরা যাতে প্রাণীগুলোর সাথে বেশ চমকপ্রদভাবে পরিচিত হতে পারে, সেই ব্যবস্থা পাকা হয়ে ওঠে।[১৮০] উল্টোদিক থেকে ডাইনোসরদের জনপ্রিয়তাও যে তাদের সম্পর্কে বৈজ্ঞানিক আবিষ্কারের একটি গুরুত্বপূর্ণ চালিকাশক্তি সে'কথা অস্বীকার করা যায় না। বেশ কিছু নমুনার আবিষ্কারের পিছনে এই জনপ্রিয়তার প্রত্যক্ষ বা পরোক্ষ সংযোগ আছে। যেমন, ১৮৮০-৯০ এর দশক নাগাদ মার্কিন যুক্তরাষ্ট্রের বিভিন্ন জাদুঘর তাদের ডাইনোসর সংগ্রহ বাড়ানোর তাগিদে বিজ্ঞানী কোপ ও মার্শের জীবাশ্ম যুদ্ধে মদত দিয়েছিল। গোটা প্রক্রিয়াটা ডাইনোসর গবেষণার জগতের প্রভূত উপকার সাধন করেছিল।[১৮১]
ডাইনোসরদের জনপ্রিয়তা সাহিত্য, চলচ্চিত্র এবং অন্যান্য গণমাধ্যমে তাদের প্রবেশ নিশ্চিত করেছে। ১৮৫২ খ্রিঃ চার্লস ডিকেন্স তার উপন্যাস ব্লিক হাউসে একবার আলগোছে ডাইনোসরদের উল্লেখ[১৮২] করেছিলেন। এর পর একাধিক গল্পে তারা অনেক বেশি গুরুত্বপূর্ণ ভূমিকা নিয়েছে। জুল ভার্নের ১৮৬৪ খ্রিঃ প্রকাশিত উপন্যাস জার্নি টু দ্য সেন্টার অফ দ্য আর্থ, স্যার আর্থার কোনান ডয়েলের ১৯১২ খ্রিঃ প্রকাশিত বই দ্য লস্ট ওয়ার্ল্ড, ১৯৩৩ খ্রিঃ এর বিখ্যাত চলচ্চিত্র কিং কং, ১৯৫৪ খ্রিঃ মুক্তিপ্রাপ্ত গডজিলা ও তার অজস্র অনুসারী চলচ্চিত্র (সিকোয়েল), ১৯৯০ খ্রিঃ মাইকেল ক্রিকটনের লেখা কালজয়ী উপন্যাস জুরাসিক পার্ক এবং তার ভিত্তিতে নির্মিত ১৯৯৩ খ্রিঃ মুক্তিপ্রাপ্ত চলচ্চিত্রটি — এগুলো হল ডাইনোসরদের উল্লেখযোগ্য কাল্পনিক পুনর্নির্মাণের মধ্যে মাত্র কয়েকটা। বিজ্ঞান বিষয়ক নিবন্ধের লেখক পুরাজীববিদ ও অন্যান্য বিজ্ঞানী বা বিজ্ঞানানুরাগী ব্যক্তিরা প্রায়ই বিজ্ঞানের বিভিন্ন নিয়ম সম্বন্ধে সাধারণ মানুষকে সচেতন করার জন্য ডাইনোসরদের একটি উপায় হিসেবে ব্যবহার করে থাকেন, অর্থাৎ জনপ্রিয় কোনো বিষয় নিয়ে কথা বললে জনগণ সেটা মনোযোগ দিয়ে শুনবে — এটা হল অনুসিদ্ধান্ত। বিজ্ঞাপনের জগতে ডাইনোসরেরা প্রায়ই আবির্ভূত হয়। অনেক কোম্পানি নানা ক্ষেত্রে তাদের প্রতীক হিসেবে ব্যবহার করে থাকে; হয় তাদের নিজেদের পণ্যকে উৎকৃষ্ট বোঝাতে বা তাদের প্রতিদ্বন্দ্বীদের অলস, মূর্খ বা প্রাচীনপন্থী প্রতিপন্ন করতে।[১৮৩]
আরও দেখুন
সম্পাদনাপাদটীকা এবং তথ্যসূত্র
সম্পাদনা- ↑ ক খ গ Rey LV, Holtz, Jr TR (২০০৭)। Dinosaurs: the most complete, up-to-date encyclopedia for dinosaur lovers of all ages। New York: Random House। আইএসবিএন 0-375-82419-7। উদ্ধৃতি ত্রুটি:
<ref>ট্যাগ বৈধ নয়; আলাদা বিষয়বস্তুর সঙ্গে "Holtz2008" নামটি একাধিক বার সংজ্ঞায়িত করা হয়েছে - ↑ ক খ Feduccia, A. (২০০২)। "Birds are dinosaurs: simple answer to a complex problem"। The Auk। 119 (4): 1187–1201। ডিওআই:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2।
- ↑ Wang, S.C., and Dodson, P. (২০০৬)। "Estimating the Diversity of Dinosaurs"। Proceedings of the National Academy of Sciences USA। 103 (37): 13601–13605। ডিওআই:10.1073/pnas.0606028103। পিএমআইডি 16954187। পিএমসি 1564218
 । বিবকোড:2006PNAS..10313601W।
। বিবকোড:2006PNAS..10313601W।
- ↑ Amos J (২০০৮-০৯-১৭)। "Will the real dinosaurs stand up?"। BBC News। সংগ্রহের তারিখ ২০১১-০৩-২৩।
- ↑ ক খ MacLeod, N, Rawson, PF, Forey, PL, Banner, FT, Boudagher-Fadel, MK, Bown, PR, Burnett, JA, Chambers, P, Culver, S, Evans, SE, Jeffery, C, Kaminski, MA, Lord, AR, Milner, AC, Milner, AR, Morris, N, Owen, E, Rosen, BR, Smith, AB, Taylor, PD, Urquhart, E & Young, JR (১৯৯৭)। "The Cretaceous–Tertiary biotic transition"। Journal of the Geological Society। 154 (2): 265–292। ডিওআই:10.1144/gsjgs.154.2.0265। উদ্ধৃতি ত্রুটি:
<ref>ট্যাগ বৈধ নয়; আলাদা বিষয়বস্তুর সঙ্গে "MacLeod" নামটি একাধিক বার সংজ্ঞায়িত করা হয়েছে - ↑ ক খ গ Carpenter, Kenneth (২০০৬)। "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus"। Foster, John R.; and Lucas, Spencer G. (eds.)। Paleontology and Geology of the Upper Jurassic Morrison Formation। New Mexico Museum of Natural History and Science Bulletin 36। Albuquerque: New Mexico Museum of Natural History and Science। পৃষ্ঠা 131–138।
- ↑ ক খ Owen, R (১৮৪২)। Report on British Fossil Reptiles." Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841। London: John Murray। পৃষ্ঠা 60–204।
- ↑ "Liddell–Scott–Jones Lexicon of Classical Greek"। সংগ্রহের তারিখ ২০০৮-০৮-০৫।
- ↑ Farlow, J.O., and Brett-Surman, M.K. (১৯৯৭)। "Preface"। Farlow, J.O., and Brett-Surman, M.K. (eds.)। The Complete Dinosaur। Indiana University Press। পৃষ্ঠা ix–xi। আইএসবিএন 0-253-33349-0।
- ↑ ক খ Benton, Michael J. (২০০৪)। "Origin and relationships of Dinosauria"। Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.)। The Dinosauria (2nd সংস্করণ)। Berkeley: University of California Press। পৃষ্ঠা 7–19। আইএসবিএন 0-520-24209-2।
- ↑ Olshevsky, G. (২০০০)। "An annotated checklist of dinosaur species by continent"। Mesozoic Meanderings। 3: 1–157।
- ↑ ক খ Langer, Max C.; Martin D. Ezcurra; Jonathas S. Bittencourt; Fernando E. Novas (২০১০)। "The origin and early evolution of dinosaurs" (পিডিএফ)। Biological Reviews। 85 (1): 65–66, 82। ২৯ অক্টোবর ২০১৩ তারিখে মূল (পিডিএফ) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ৭ জুন ২০১৪। উদ্ধৃতি ত্রুটি:
<ref>ট্যাগ বৈধ নয়; আলাদা বিষয়বস্তুর সঙ্গে "Letal05" নামটি একাধিক বার সংজ্ঞায়িত করা হয়েছে - ↑ Padian, K. (2004). "Basal Avialae". In Weishampel, D.B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. আইএসবিএন ০-৫২০-২৪২০৯-২
- ↑ Glut, Donald F. (১৯৯৭)। Dinosaurs: The Encyclopedia। Jefferson, North Carolina: McFarland & Co। পৃষ্ঠা 40। আইএসবিএন 0-89950-917-7। উদ্ধৃতি ত্রুটি:
<ref>ট্যাগ বৈধ নয়; আলাদা বিষয়বস্তুর সঙ্গে "DFG97" নামটি একাধিক বার সংজ্ঞায়িত করা হয়েছে - ↑ Lambert, David; the Diagram Group (১৯৯০)। The Dinosaur Data Book। New York: Avon Books। পৃষ্ঠা 288। আইএসবিএন 0-380-75896-2। উদ্ধৃতি ত্রুটি:
<ref>ট্যাগ বৈধ নয়; আলাদা বিষয়বস্তুর সঙ্গে "DL90" নামটি একাধিক বার সংজ্ঞায়িত করা হয়েছে - ↑ Morales, Michael (১৯৯৭)। "Nondinosaurian vertebrates of the Mesozoic"। Farlow JO, Brett-Surman MK। The Complete Dinosaur। Bloomington: Indiana University Press। পৃষ্ঠা 607–624। আইএসবিএন 0-253-33349-0।
- ↑ Wang, S.C., and Dodson, P. (2006). "Estimating the Diversity of Dinosaurs". Proceedings of the National Academy of Sciences USA 103 (37): 13601–13605. Bibcode:2006PNAS..10313601W. doi:10.1073/pnas.0606028103. PMC 1564218. PMID 16954187.
- ↑ Russell, Dale A. (1995). "China and the lost worlds of the dinosaurian era". Historical Biology 10: 3–12. doi:10.1080/10292389509380510.
- ↑ Amos J (2008-09-17). "Will the real dinosaurs stand up?". BBC News. Retrieved 2011-03-23.
- ↑ MacLeod, N, Rawson, PF, Forey, PL, Banner, FT, Boudagher-Fadel, MK, Bown, PR, Burnett, JA, Chambers, P, Culver, S, Evans, SE, Jeffery, C, Kaminski, MA, Lord, AR, Milner, AC, Milner, AR, Morris, N, Owen, E, Rosen, BR, Smith, AB, Taylor, PD, Urquhart, E & Young, JR (1997). "The Cretaceous–Tertiary biotic transition". Journal of the Geological Society 154 (2): 265–292. doi:10.1144/gsjgs.154.2.0265.
- ↑ Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; and Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology 38 (2): 139–142. doi:10.1130/G30402.1.
- ↑ Brusatte, Stephen L. (2012). Dinosaur Paleobiology (1. ed.). New York: Wiley, J. pp. 9–20, 21. আইএসবিএন ৯৭৮-০-৪৭০-৬৫৬৫৮-৭.
- ↑ Nesbitt S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades". Bulletin of the American Museum of Natural History 352: 1–292. doi:10.1206/352.1.
- ↑ Benton, Michael J. (2004). "Origin and relationships of Dinosauria". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 7–19. আইএসবিএন ০-৫২০-২৪২০৯-২
- ↑ This was recognized not later than 1909: "Dr. Holland and the Sprawling Sauropods". Archived from the original on 2011-06-12. The arguments and many of the images are also presented in Desmond, A. (1976). Hot Blooded Dinosaurs. DoubleDay. আইএসবিএন ০-৩৮৫-২৭০৬৩-১.
- ↑ Benton, M.J. (2004). Vertebrate Paleontology. Blackwell Publishers. xii–452. আইএসবিএন ০-৬৩২-০৫৬১৪-২.
- ↑ Cowen, Richard (2004). "Dinosaurs". History of Life (4th ed.). Blackwell Publishing. pp. 151–175. আইএসবিএন ১-৪০৫১-১৭৫৬-৭. OCLC 53970577.
- ↑ ক খ Kubo, T.; Benton, Michael J. (2007). "Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates". Palaeontology 50 (6): 1519–1529. doi:10.1111/j.1475-4983.2007.00723.x.
- ↑ Kump LR, Pavlov A & Arthur MA (২০০৫)। "Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia"। Geology। 33 (5): 397–400। ডিওআই:10.1130/G21295.1। বিবকোড:2005Geo....33..397K।
- ↑ Tanner LH, Lucas SG & Chapman MG (২০০৪)। "Assessing the record and causes of Late Triassic extinctions" (পিডিএফ)। Earth-Science Reviews। 65 (1–2): 103–139। ডিওআই:10.1016/S0012-8252(03)00082-5। বিবকোড:2004ESRv...65..103T। অক্টোবর ২৫, ২০০৭ তারিখে মূল (PDF) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০০৭-১০-২২।
- ↑ অ্যালকোবার, অস্কার এ.; মার্টিনেজ, রিকার্ডো এন. (২০১০)। "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina"। ZooKeys। ৬৩ (৬৩): ৫৫–৮১। ডিওআই:10.3897/zookeys.63.550। পিএমআইডি 21594020। পিএমসি 3088398 । [১][স্থায়ীভাবে অকার্যকর সংযোগ]
- ↑ ক খ Sereno PC (১৯৯৯)। "The evolution of dinosaurs"। Science। 284 (5423): 2137–2147। ডিওআই:10.1126/science.284.5423.2137। পিএমআইডি 10381873।
- ↑ Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (১৯৯৩)। "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria"। Nature। 361 (6407): 64–66। ডিওআই:10.1038/361064a0। বিবকোড:1993Natur.361...64S।
- ↑ Nesbitt, S. J., Barrett, P. M., Werning, S., Sidor, C. A., and A. J. Charig. (২০১২). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania." Biology Letters.
- ↑ Justin A. Spielmann, Spencer G. Lucas and Adrian P. Hunt (২০১৩)। "The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A." (পিডিএফ)। New Mexico Museum of Natural History and Science Bulletin। ৬১: ৫৬২–৫৬৬। ১ অক্টোবর ২০১৫ তারিখে মূল (পিডিএফ) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ১০ জুন ২০১৪।
- ↑ Jerzy Dzik, Tomasz Sulej, and Grzegorz Niedźwiedzki (২০০৮)। "A dicynodont−theropod association in the latest Triassic of Poland"। Acta Palaeontologica Polonica। ৫৩ (৪): ৭৩৩–৭৩৮।
- ↑ ক খ গ ঘ Holtz, Thomas R., Jr. (২০০৪)। "Mesozoic biogeography of Dinosauria"। Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.)। The Dinosauria (2nd সংস্করণ)। Berkeley: University of California Press। পৃষ্ঠা 627–642। আইএসবিএন 0-520-24209-2। অজানা প্যারামিটার
|coauthors=উপেক্ষা করা হয়েছে (|author=ব্যবহারের পরামর্শ দেয়া হচ্ছে) (সাহায্য) - ↑ ক খ গ ঘ ঙ Fastovsky, David E.; Smith, Joshua B. (২০০৪)। "Dinosaur paleoecology"। Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka। The Dinosauria (2nd সংস্করণ)। Berkeley: University of California Press। পৃষ্ঠা 614–626। আইএসবিএন 0-520-24209-2।
- ↑ ক খ গ ঘ ঙ চ Padian K (২০০৪)। "Basal avialae"। Weishampel DB, Dodson P, Osmólska H। The Dinosauria (2d edition)। University of California Press। পৃষ্ঠা 210–231। আইএসবিএন 0-520-24209-2।
- ↑ Sereno, P.C.; Wilson, JA; Witmer, LM; Whitlock, JA; Maga, A; Ide, O; Rowe, TA; Kemp, Tom (২০০৭)। Kemp, Tom, সম্পাদক। "Structural extremes in a Cretaceous dinosaur"। PLoS ONE। 2 (11): e1230। ডিওআই:10.1371/journal.pone.0001230। পিএমআইডি 18030355। পিএমসি 2077925 । বিবকোড:2007PLoSO...2.1230S।
- ↑ Prasad, V.; Strömberg, CA; Alimohammadian, H; Sahni, A (২০০৫)। "Dinosaur coprolites and the early evolution of grasses and grazers"। Science। 310 (5751): 1170–1180। ডিওআই:10.1126/science.1118806। পিএমআইডি 16293759। বিবকোড:2005Sci...310.1177P।
- ↑ Archibald, J. David; Fastovsky, David E. (২০০৪)। "Dinosaur Extinction"। Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.)। The Dinosauria (2nd সংস্করণ)। Berkeley: University of California Press। পৃষ্ঠা 672–684। আইএসবিএন 0-520-24209-2।
- ↑ Lindow, B.E.K. (2011). "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification." In Dyke, G. and Kaiser, G. (eds.)Living Dinosaurs: The Evolutionary History of Modern Birds, John Wiley & Sons, Ltd, Chichester, UK. ডিওআই:10.1002/9781119990475.ch14
- ↑ ক খ Brusatte, Stephen L. (২০১২)। Dinosaur Paleobiology (1. সংস্করণ)। New York: Wiley, J। পৃষ্ঠা 9–20, 21। আইএসবিএন 978-0-470-65658-7।
- ↑ Benton, M.J. (২০০৪)। Vertebrate Paleontology। Blackwell Publishers। xii–452। আইএসবিএন 0-632-05614-2।
- ↑ Paul, G.S. (১৯৮৮)। Predatory Dinosaurs of the World। New York: Simon and Schuster। পৃষ্ঠা 248–250। আইএসবিএন 0-671-61946-2।
- ↑ Clark J.M., Maryanska T., Barsbold R (২০০৪)। "Therizinosauroidea"। Weishampel DB, Dodson P, Osmólska H। The Dinosauria (2d edition)। University of California Press। পৃষ্ঠা 151–164। আইএসবিএন 0-520-24209-2।
- ↑ Norell MA, Makovicky PJ (২০০৪)। "Dromaeosauridae"। Weishampel DB, Dodson P, Osmólska H। The Dinosauria (2d edition)। University of California Press। পৃষ্ঠা 196–210। আইএসবিএন 0-520-24209-2।
- ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ ক খ Dal Sasso, C. and Signore, M. (১৯৯৮)। "Exceptional soft-tissue preservation in a theropod dinosaur from Italy"। Nature। 392 (6674): 383–387। ডিওআই:10.1038/32884। বিবকোড:1998Natur.392..383D।
- ↑ ক খ গ Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. (২০০৫)। "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex"। Science। 307 (5717): 1952–1955। ডিওআই:10.1126/science.1108397। পিএমআইডি 15790853। বিবকোড:2005Sci...307.1952S।
- ↑ ক খ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Weishampel, D.B., Dodson, P., Oslmolska, H. (1990). "The Dinosauria". University of California Press. pp. 733. আইএসবিএন ০-৫২০-০৬৭২৭-৪
- ↑ Farlow JA (১৯৯৩)। "On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs"। Dodson, Peter; and Gingerich, Philip। Functional Morphology and Evolution। American Journal of Science, Special Volume 293-A। পৃষ্ঠা 167–199।
- ↑ Peczkis, J. (১৯৯৪)। "Implications of body-mass estimates for dinosaurs"। Journal of Vertebrate Paleontology। 14 (4): 520–33। ডিওআই:10.1080/02724634.1995.10011575।
- ↑ "Anatomy and evolution"। National Museum of Natural History। সংগ্রহের তারিখ ২০০৭-১১-২১।
- ↑ Sander, P. Martin; Christian, Andreas; Clauss, Marcus; Fechner, Regina; Gee, Carole T.; Griebeler, Eva-Maria; Gunga, Hanns-Christian; Hummel, Jürgen; Mallison, Heinrich; ও অন্যান্য (২০১১)। "Biology of the sauropod dinosaurs: the evolution of gigantism"। Biological Reviews। 86 (1): 117–155। ডিওআই:10.1111/j.1469-185X.2010.00137.x। পিএমআইডি 21251189। পিএমসি 3045712 ।
- ↑ Paul, Gregory S. (২০১০)। Princeton Field Guide to Dinosaurs। Princeton University Press। আইএসবিএন 978-0-691-13720-9।
- ↑ Colbert, Edwin Harris (১৯৭১)। Men and dinosaurs: the search in field and laboratory। Harmondsworth [Eng.]: Penguin। আইএসবিএন 0-14-021288-4।
- ↑ Lucas, H., Hecket, H. & S. (2004). "Reappraisal of Seismosaurus, a Late Jurassic Sauropod". Proceeding, Annual Meeting of the Society of Paleontology 36 (5): 422.
- ↑ Lovelace, David M. (২০০৭)। "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny"। Arquivos do Museu Nacional। 65 (4): 527–544।
- ↑ dal Sasso C, Maganuco S, Buffetaut E, Mendez MA (২০০৬)। "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities" (পিডিএফ)। Journal of Vertebrate Paleontology। 25 (4): 888–896। ডিওআই:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2। ২০১১-০৪-২৯ তারিখে মূল (PDF) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০১১-০৫-০৫।
- ↑ Therrien, F.; Henderson, D.M. (২০০৭)। "My theropod is bigger than yours ... or not: estimating body size from skull length in theropods"। Journal of Vertebrate Paleontology। 27 (1): 108–115। ডিওআই:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2।
- ↑ ক খ Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (২০০৮)। "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers"। Nature। 455 (7216): 1105–1108। ডিওআই:10.1038/nature07447। পিএমআইডি 18948955। বিবকোড:2008Natur.455.1105Z।
- ↑ ক খ Xu X, Zhao Q, Norell M, Sullivan C, Hone D, Erickson G, Wang XL, Han FL, Guo Y (২০০৮)। "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin"। Chinese Science Bulletin। 54 (3): 430–435। ডিওআই:10.1007/s11434-009-0009-6।
- ↑ Butler, R.J.; Zhao, Q. (২০০৯)। "The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus yansiensis from the Late Cretaceous of China"। Cretaceous Research। 30 (1): 63–77। ডিওআই:10.1016/j.cretres.2008.03.002।
- ↑ Yans J, Dejax J, Pons D, Dupuis C & Taquet P (২০০৫)। "Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)"। Comptes Rendus Palevol (French ভাষায়)। 4 (1–2): 135–150। ডিওআই:10.1016/j.crpv.2004.12.003।
- ↑ Day, J.J.; Upchurch, P; Norman, DB; Gale, AS; Powell, HP (২০০২)। "Sauropod trackways, evolution, and behavior"। Science। 296 (5573): 1659। ডিওআই:10.1126/science.1070167। পিএমআইডি 12040187।
- ↑ Wright, Joanna L. (২০০৫)। "Steps in understanding sauropod biology"। Curry Rogers, Kristina A.; and Wilson, Jeffrey A.। The Sauropods: Evolution and Paleobiology। Berkeley: University of California Press। পৃষ্ঠা 252–284। আইএসবিএন 0-520-24623-3।
- ↑ Varricchio, D.J.; Sereno, Paul C.; Xijin, Zhao; Lin, Tan; Wilson, Jeffery A.; Lyon, Gabrielle H. (২০০৮)। "Mud-trapped herd captures evidence of distinctive dinosaur sociality" (PDF)। Acta Palaeontologica Polonica। 53 (4): 567–578। ডিওআই:10.4202/app.2008.0402। সংগ্রহের তারিখ ২০১১-০৫-০৬।
- ↑ Lessem, Don; Glut, Donald F. (১৯৯৩)। "Allosaurus"। The Dinosaur Society's Dinosaur Encyclopedia। Random House। পৃষ্ঠা 19–20। আইএসবিএন 0-679-41770-2।
- ↑ Maxwell, W. D.; Ostrom, John (১৯৯৫)। "Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations"। Journal of Vertebrate Paleontology। 15 (4): 707–712। ডিওআই:10.1080/02724634.1995.10011256।
- ↑ Roach, Brian T.; Brinkman, Daniel L. (২০০৭)। "A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs"। Bulletin of the Peabody Museum of Natural History। 48 (1): 103–138। ডিওআই:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2।
- ↑ Tanke, Darren H. (১৯৯৮)। "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (পিডিএফ)। Gaia (15): 167–184। ২০০৮-০২-২৭ তারিখে মূল (PDF) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০১৪-০৬-০৭।
- ↑ "The Fighting Dinosaurs"। American Museum of Natural History। ২০১২-০১-১৮ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০০৭-১২-০৫।
- ↑ ক খ Carpenter, K. (১৯৯৮)। "Evidence of predatory behavior by theropod dinosaurs"। Gaia। 15: 135–144।[স্থায়ীভাবে অকার্যকর সংযোগ]
- ↑ Rogers, Raymond R.; Krause, DW; Curry Rogers, K (২০০৭)। "Cannibalism in the Madagascan dinosaur Majungatholus atopus"। Nature। 422 (6931): 515–518। ডিওআই:10.1038/nature01532। পিএমআইডি 12673249।
- ↑ Schmitz, L.; Motani, R. (২০১১)। "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology"। Science। 332 (6030): 705–708। ডিওআই:10.1126/science.1200043। পিএমআইডি 21493820। বিবকোড:2011Sci...332..705S।
- ↑ Varricchio DJ, Martin, AJ and Katsura, Y (২০০৭)। "First trace and body fossil evidence of a burrowing, denning dinosaur"। Proceedings of the Royal Society B। 274 (1616): 1361–1368। ডিওআই:10.1098/rspb.2006.0443। পিএমআইডি 17374596। পিএমসি 2176205 ।
- ↑ Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. আইএসবিএন ০-৫২০-২০০৯৪-২
- ↑ Alexander RM (২০০৬)। "Dinosaur biomechanics"। Proceedings of the Royal Society B। 273 (1596): 1849–1855। ডিওআই:10.1098/rspb.2006.3532। পিএমআইডি 16822743। পিএমসি 1634776 ।
- ↑ Goriely A & McMillen T (২০০২)। "Shape of a cracking whip"। Physical Review Letters। 88 (24): 244301। ডিওআই:10.1103/PhysRevLett.88.244301। পিএমআইডি 12059302। বিবকোড:2002PhRvL..88x4301G।
- ↑ Henderson, D.M. (২০০৩)। "Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis"। Canadian Journal of Zoology। 81 (8): 1346–1357। ডিওআই:10.1139/z03-122।
- ↑ ক খ গ ঘ Senter, P. (২০০৮)। "Voices of the past: a review of Paleozoic and Mesozoic animal sounds"। Historical Biology। 20 (4): 255–287। ডিওআই:10.1080/08912960903033327।
- ↑ Hopson, James A. (১৯৭৫)। "The evolution of cranial display structures in hadrosaurian dinosaurs"। Paleobiology। 1 (1): 21–43।
- ↑ Diegert, Carl F. (১৯৯৮)। "A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen"। Journal of Vertebrate Paleontology। 18 (3, Suppl.): 38A।
- ↑ Currie, Philip J and Kevin Padian, সম্পাদক (১৯৯৭)। Encyclopedia of Dinosaurs। Academic Press। পৃষ্ঠা 206।
- ↑ Hansell M (2000). Bird Nests and Construction Behaviour. University of Cambridge Press আইএসবিএন ০-৫২১-৪৬০৩৮-৭
- ↑ Varricchio, David J.; Horner, John J.; Jackson, Frankie D. (২০০২)। "Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus"। Journal of Vertebrate Paleontology। 22 (3): 564–576। ডিওআই:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2।
- ↑ Lee, Andrew H.; Werning, S (২০০৮)। "Sexual maturity in growing dinosaurs does not fit reptilian growth models"। Proceedings of the National Academy of Sciences। 105 (2): 582–587। ডিওআই:10.1073/pnas.0708903105। পিএমআইডি 18195356। পিএমসি 2206579 । বিবকোড:2008PNAS..105..582L। ১৩ মে ২০০৮ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ৭ জুন ২০১৪।
- ↑ Horner, J.R.; Makela, Robert (১৯৭৯)। "Nest of juveniles provides evidence of family structure among dinosaurs"। Nature। 282 (5736): 296–298। ডিওআই:10.1038/282296a0। বিবকোড:1979Natur.282..296H।
- ↑ Chiappe, Luis M. (২০০৫)। "Nesting titanosaurs from Auca Mahuevo and adjacent sites"। Curry Rogers, Kristina A.; and Wilson, Jeffrey A.। The Sauropods: Evolution and Paleobiology। Berkeley: University of California Press। পৃষ্ঠা 285–302। আইএসবিএন 0-520-24623-3। অজানা প্যারামিটার
|coauthors=উপেক্ষা করা হয়েছে (|author=ব্যবহারের পরামর্শ দেয়া হচ্ছে) (সাহায্য) - ↑ "Discovering Dinosaur Behavior: 1960–present view"। Encyclopædia Britannica। সংগ্রহের তারিখ ২০১১-০৫-০৫।
- ↑ Hopp, Thomas P.; Mark J. Orsen (২০০৪)। "11: Dinosaur Brooding Behavior and the Origin of Flight Feathers"। Philip J. Currie, Eva B. Koppelhus, Martin A. Shugar, Joanna L. Wright, James O. Farlow। Feathered Dragons: Studies on the Transition from Dinosaurs to Birds (পিডিএফ)। Bloomington: Indiana University Press। পৃষ্ঠা 234–250। সংগ্রহের তারিখ ১০ ডিসেম্বর ২০১৩।
- ↑ Reisz RR, Scott, D Sues, H-D, Evans, DC & Raath, MA (২০০৫)। "Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance"। Science। 309 (5735): 761–764। ডিওআই:10.1126/science.1114942। পিএমআইডি 16051793। বিবকোড:2005Sci...309..761R।
- ↑ Clark NDL, Booth P, Booth CL, Ross DA (২০০৪)। "Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye" (PDF)। Scottish Journal of Geology। 40 (1): 13–21। ডিওআই:10.1144/sjg40010013। সংগ্রহের তারিখ ২০১১-০৫-০৫।[স্থায়ীভাবে অকার্যকর সংযোগ]
- ↑ Horner, J. & Gorman, J. (1988). Digging Dinosaurs: The Search that Unraveled the Mystery of Baby Dinosaurs. Workman Publishing Co.
- ↑ Chinsamy A, Hillenius WJ (২০০৪)। "Physiology of nonavian dinosaurs"। Weishampel DB, Dodson P, Osmólska H। The Dinosauria (2nd সংস্করণ)। University of California Press। পৃষ্ঠা 643–659। আইএসবিএন 0-520-24209-2।
- ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ "Hot-Blooded or Cold-Blooded??"। University of California Museum of Paleontology। জানুয়ারি ২৫, ২০১২ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ফেব্রুয়ারি ১২, ২০১২।
- ↑ Parsons, Keith M. (২০০১)। Drawing out Leviathan: Dinosaurs and the science wars। Bloomington: Indiana University Press। পৃষ্ঠা 22–48। আইএসবিএন 0-253-33937-5।
- ↑ Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA; ও অন্যান্য (২০০৮)। Kemp, Tom, সম্পাদক। "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina"। PLoS ONE। 3 (9): e3303doi=10.1371/journal.pone.0003303। ডিওআই:10.1371/journal.pone.0003303। পিএমআইডি 18825273। পিএমসি 2553519 । বিবকোড:2008PLoSO...3.3303S। সংগ্রহের তারিখ ২০০৮-০৯-২৯। অজানা প্যারামিটার
|month=উপেক্ষা করা হয়েছে (সাহায্য) - ↑ Reid, R.E.H. (১৯৯৭)। "Dinosaurian Physiology: the Case for "Intermediate" Dinosaurs"। Farlow, J.O., and Brett-Surman, M.K.। The Complete Dinosaur। Bloomington: Indiana University Press। পৃষ্ঠা 449–473। আইএসবিএন 0-253-33349-0।
- ↑ Ehrlich, Paul R.; David S. Dobkin; Darryl Wheye (১৯৮৮)। "Drinking"। Birds of Stanford। Standford University। সংগ্রহের তারিখ ২০০৭-১২-১৩।
- ↑ Tsahar, Ella; Martínez Del Rio, C; Izhaki, I; Arad, Z (২০০৫)। "Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores" (Free full text)। Journal of Experimental Biology। 208 (6): 1025–34। ডিওআই:10.1242/jeb.01495। পিএমআইডি 15767304।[স্থায়ীভাবে অকার্যকর সংযোগ]
- ↑ Skadhauge, E; Erlwanger, KH; Ruziwa, SD; Dantzer, V; Elbrønd, VS; Chamunorwa, JP (২০০৩)। "Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?"। Comparative biochemistry and physiology. Part A, Molecular & integrative physiology। 134 (4): 749–755। ডিওআই:10.1016/S1095-6433(03)00006-0। পিএমআইডি 12814783।
- ↑ Preest, Marion R.; Beuchat, Carol A. (এপ্রিল ১৯৯৭)। "Ammonia excretion by hummingbirds"। Nature। 386 (6625): 561–62। ডিওআই:10.1038/386561a0। বিবকোড:1997Natur.386..561P।
- ↑ Mora, J.; Martuscelli, J; Ortiz Pineda, J; Soberon, G (জুলাই ১৯৬৫)। "The Regulation of Urea-Biosynthesis Enzymes in Vertebrates" (PDF)। Biochemical Journal। 96 (1): 28–35। পিএমআইডি 14343146। পিএমসি 1206904 ।
- ↑ Packard, Gary C. (১৯৬৬)। "The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates"। The American Naturalist। 100 (916): 667–82। জেস্টোর 2459303। ডিওআই:10.1086/282459।
- ↑ Balgooyen, Thomas G. (১ অক্টোবর ১৯৭১)। "Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius)" (PDF)। Condor। 73 (3): 382–85। জেস্টোর 1365774। ডিওআই:10.2307/1365774।
- ↑ Huxley, Thomas H. (১৮৬৮)। "On the animals which are most nearly intermediate between birds and reptiles"। Annals of the Magazine of Natural History। 4 (2): 66–75।
- ↑ Heilmann, Gerhard (১৯২৬)। The Origin of Birds। London: Witherby। পৃষ্ঠা 208pp। আইএসবিএন 0-486-22784-7।
- ↑ Osborn, Henry Fairfield (১৯২৪)। "Three new Theropoda, Protoceratops zone, central Mongolia" (পিডিএফ)। American Museum Novitates। 144: 1–12। ২৭ মার্চ ২০০৯ তারিখে মূল (PDF) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ৭ জুন ২০১৪।
- ↑ Ostrom, John H. (১৯৭৩)। "The ancestry of birds"। Nature। 242 (5393): 136। ডিওআই:10.1038/242136a0। বিবকোড:1973Natur.242..136O।
- ↑ Holtz, Jr., T.R. (২০০০)। "Classification and evolution of the dinosaur groups"। Paul, G.S.। The Scientific American Book of Dinosaurs। St. Martin's Press। পৃষ্ঠা 140–168। আইএসবিএন 0-312-26226-4।
- ↑ Gauthier, Jacques. (১৯৮৬)। "Saurischian monophyly and the origin of birds"। Padian, Kevin. (ed.)। The Origin of Birds and the Evolution of Flight। Memoirs of the California Academy of Sciences 8। পৃষ্ঠা 1–55।
- ↑ Mayr, G., Pohl, B. and Peters, D.S. (২০০৫)। "A Well-Preserved Archaeopteryx Specimen with Theropod Features"। Science। 310 (5753): 1483–1486। ডিওআই:10.1126/science.1120331। পিএমআইডি 16322455। বিবকোড:2005Sci...310.1483M।
- ↑ ক খ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Godefroit, P., Sinitsa, S., Dhouailly, D., Bolotsky, Y., and Sizov, A. "Feather-like structures and scales in a Jurassic neornithischian dinosaur from Siberia." Program and Abstracts of the 73rd Meeting of the Society of Vertebrate Paleontology ওয়েব্যাক মেশিনে আর্কাইভকৃত ১৯ অক্টোবর ২০১৩ তারিখে, October 2013.
- ↑ Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; and Jia C. (২০০৪)। "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids"। Nature। 431 (7009): 680–684। ডিওআই:10.1038/nature02855। পিএমআইডি 15470426। বিবকোড:2004Natur.431..680X।
- ↑ Göhlich, U.B.; Chiappe, LM (২০০৬)। "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago"। Nature। 440 (7082): 329–332। ডিওআই:10.1038/nature04579। পিএমআইডি 16541071। বিবকোড:2006Natur.440..329G।
- ↑ Kellner, A.W.A., Wang, X., Tischlinger, H., Campos, D., Hone, D.W.E. and Meng, X. (2009). "The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane." Proceedings of the Royal Society B, published online before print August 5, 2009, ডিওআই:10.1098/rspb.2009.0846
- ↑ Wellnhofer, P (১৯৮৮)। "Ein neuer Exemplar von Archaeopteryx"। Archaeopteryx। 6: 1–30।
- ↑ ক খ Schweitzer, M.H.; Watt, J.A.; Avci, R.; Knapp, L.; Chiappe, L.; Norell, M.; Marshall, M. (১৯৯৯)। "Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid,Shuvuuia deserti"। Journal of Experimental Zoology। 285 (2): 146–57। ডিওআই:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A। পিএমআইডি 10440726।
- ↑ Lingham-Soliar, T. (২০০৩)। "The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers"। Naturwissenschaften। 90 (12): 563–567। ডিওআই:10.1007/s00114-003-0483-7। পিএমআইডি 14676953। বিবকোড:2003NW.....90..563L।
- ↑ Feduccia, A.; Lingham-Soliar, T; Hinchliffe, JR (২০০৫)। "Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence"। Journal of Morphology। 266 (2): 125–166। ডিওআই:10.1002/jmor.10382। পিএমআইডি 16217748।
- ↑ Lingham-Soliar, T.; Feduccia, A; Wang, X (২০০৭)। "A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres"। Proceedings of the Royal Society B। 274 (1620): 1823–9। ডিওআই:10.1098/rspb.2007.0352। পিএমআইডি 17521978। পিএমসি 2270928 ।
- ↑ Prum, Richard O. (এপ্রিল ২০০৩)। "Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002"। The Auk। 120 (2): 550–61। জেস্টোর 4090212। ডিওআই:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2।
- ↑ Archaeopteryx: a missing link. Berkeley: University of California. Museum of Paleontology.
- ↑ O'Connor, P.M. & Claessens, L.P.A.M. (২০০৫)। "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs"। Nature। 436 (7048): 253–256। ডিওআই:10.1038/nature03716। পিএমআইডি 16015329। বিবকোড:2005Natur.436..253O।
- ↑ Sereno, P.C.; Martinez, RN; Wilson, JA; Varricchio, DJ; Alcober, OA; Larsson, HC; Kemp, Tom (সেপ্টেম্বর ২০০৮)। Kemp, Tom, সম্পাদক। "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina"। PLoS ONE। 3 (9): e3303। ডিওআই:10.1371/journal.pone.0003303। পিএমআইডি 18825273। পিএমসি 2553519 । বিবকোড:2008PLoSO...3.3303S। সংগ্রহের তারিখ ২০০৮-১০-২৭।
- ↑ "Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System"। সংগ্রহের তারিখ ২০১১-০৫-০৫।
- ↑ Xu, X. and Norell, M.A. (২০০৪)। "A new troodontid dinosaur from China with avian-like sleeping posture"। Nature। 431 (7010): 838–841। ডিওআই:10.1038/nature02898। পিএমআইডি 15483610। বিবকোড:2004Natur.431..838X।
- ↑ Norell M.A., Clark J.M., Chiappe L.M., Dashzeveg D. (১৯৯৫)। "A nesting dinosaur"। Nature। 378 (6559): 774–776। ডিওআই:10.1038/378774a0। বিবকোড:1995Natur.378..774N।
- ↑ Varricchio, D. J.; Moore, J. R.; Erickson, G. M.; Norell, M. A.; Jackson, F. D.; Borkowski, J. J. (২০০৮)। "Avian Paternal Care Had Dinosaur Origin"। Science। 322 (5909): 1826–8। ডিওআই:10.1126/science.1163245। পিএমআইডি 19095938। বিবকোড:2008Sci...322.1826V।
- ↑ Wings O (২০০৭)। "A review of gastrolith function with implications for fossil vertebrates and a revised classification" (PDF)। Palaeontologica Polonica। 52 (1): 1–16। সংগ্রহের তারিখ ২০১১-০৫-০৫।
- ↑ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction – Dinosaur Evolution and the Origin of Birds. New York: W. H. Freeman.
- ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Mullen, L. (2004). "Multiple Impacts ওয়েব্যাক মেশিনে আর্কাইভকৃত ৬ এপ্রিল ২০১২ তারিখে". Astrobiology Magazine.
- ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Paul, Gregory S. (২০০২)। Dinosaurs of the air: the evolution and loss of flight in dinosaurs and birds। Johns Hopkins University Press। পৃষ্ঠা 397। আইএসবিএন 0-8018-6763-0।
- ↑ । অজানা প্যারামিটার
|1=উপেক্ষা করা হয়েছে (সাহায্য);|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV (১৯৮০)। "Extraterrestrial cause for the Cretaceous–Tertiary extinction"। Science। 208 (4448): 1095–1108। ডিওআই:10.1126/science.208.4448.1095। পিএমআইডি 17783054। বিবকোড:1980Sci...208.1095A।
- ↑ Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. (সেপ্টেম্বর ১৯৯১)। "Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico"। Geology। 19 (9): 867–871। ডিওআই:10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2। বিবকোড:1991Geo....19..867H।
- ↑ Pope KO, Ocampo AC, Kinsland GL, Smith R (১৯৯৬)। "Surface expression of the Chicxulub crater"। Geology। 24 (6): 527–30। ডিওআই:10.1130/0091-7613(1996)024<0527:SEOTCC>2.3.CO;2। পিএমআইডি 11539331। বিবকোড:1996Geo....24..527P।
- ↑ Robertson, D.S.; ও অন্যান্য (৩০ সেপ্টেম্বর ২০০৩)। "Survival in the first hours of the Cenozoic" (পিডিএফ)। Geological Society of America Bulletin। 116 (5/6): 760–768। ডিওআই:10.1130/B25402.1। বিবকোড:2004GSAB..116..760R। ১৮ সেপ্টেম্বর ২০১২ তারিখে মূল (পিডিএফ) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ১৫ জুন ২০১১।
- ↑ ক খ Hofman, C, Féraud, G & Courtillot, V (২০০০)। "40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps"। Earth and Planetary Science Letters। 180: 13–27। ডিওআই:10.1016/S0012-821X(00)00159-X। বিবকোড:2000E&PSL.180...13H।
- ↑ ক খ গ Duncan, RA & Pyle, DG (১৯৮৮)। "Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary"। Nature। 333 (6176): 841–843। ডিওআই:10.1038/333841a0। বিবকোড:1988Natur.333..841D।
- ↑ Alvarez, W (১৯৯৭)। T. rex and the Crater of Doom। Princeton University Press। পৃষ্ঠা 130–146। আইএসবিএন 978-0-691-01630-6।
- ↑ Fassett, JE, Lucas, SG, Zielinski, RA, and Budahn, JR; Lucas; Zielinski; Budahn (২০০১)। "Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA" (PDF)। Catastrophic events and mass extinctions, Lunar and Planetary Contribution। 1053: 45–46। বিবকোড:2001caev.conf.3139F। সংগ্রহের তারিখ ২০০৭-০৫-১৮।
- ↑ Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane (১৯৮৬)। "Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation"। Science। 232 (4750): 629–633। ডিওআই:10.1126/science.232.4750.629। পিএমআইডি 17781415। বিবকোড:1986Sci...232..629S।
- ↑ ক খ Fastovsky, David E.; Sheehan, Peter M. (২০০৫)। "Reply to comment on "The Extinction of the dinosaurs in North America"" (পিডিএফ)। GSA Today। 15 (7): 11। ডিওআই:10.1130/1052-5173(2005)015[11b:RTEOTD]2.0.CO;2। ২ জুন ২০১৮ তারিখে মূল (PDF) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ৭ জুন ২০১৪।
- ↑ Sullivan, RM (২০০৩)। "No Paleocene dinosaurs in the San Juan Basin, New Mexico"। Geological Society of America Abstracts with Programs। 35 (5): 15। ২০১১-০৪-০৮ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০০৭-০৭-০২।
- ↑ Fassett J.E., Heaman L.M., Simonetti A. (২০১১)। "Direct U–Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico"। Geology। 39 (2): 159–162। ডিওআই:10.1130/G31466.1।
- ↑ Dong Zhiming (১৯৯২)। Dinosaurian Faunas of China। China Ocean Press, Beijing। আইএসবিএন 3-540-52084-8। ওসিএলসি 26522845।
- ↑ "Dinosaur bones 'used as medicine'"। BBC News। ২০০৭-০৭-০৬। সংগ্রহের তারিখ ২০০৭-০৭-০৬।
- ↑ Benton, M.J. (2000). "A brief history of dinosaur paleontology". pp. 10–44, In Paul, G.S. (ed.). The Scientific American book of dinosaurs. St. Martin's Press, New York.
- ↑ ক খ Sarjeant WAS (১৯৯৭)। "The earliert discoveries"। Farlow JO, Brett-Surman MK। The Complete Dinosaur। Bloomington: Indiana University Press। পৃষ্ঠা 3–11। আইএসবিএন 0-253-33349-0।
- ↑ Gunther RT (১৯৬৮)। Life and letters of Edward Lhwyd,: Second keeper of the Museum Ashmoleanum (Early science in Oxford Volume XIV)। Dawsons of Pall Mall।
- ↑ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann:London.
- ↑ Delair, J.B.; Sarjeant, W.A.S. (২০০২)। "The earliest discoveries of dinosaurs: the records re-examined"। Proceedings of the Geologists' Association। 113 (3): 185–197। ডিওআই:10.1016/S0016-7878(02)80022-0।
- ↑ Buckland W (১৮২৪)। "Notice on the Megalosaurus or great Fossil Lizard of Stonesfield"। Transactions of the Geological Society of London। 1 (2): 390–396। ডিওআই:10.1144/transgslb.1.2.390।
- ↑ Mantell, Gideon A. (১৮২৫)। "Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex"। Philosophical Transactions of the Royal Society। 115: 179–186। জেস্টোর 107739। ডিওআই:10.1098/rstl.1825.0010।
- ↑ Sues, Hans-Dieter (১৯৯৭)। "European Dinosaur Hunters"। Farlow JO, Brett-Surman MK। The Complete Dinosaur। Bloomington: Indiana University Press। পৃষ্ঠা 14। আইএসবিএন 0-253-33349-0।
- ↑ Rupke, N. (1994). Richard Owen: A Victorian Naturalist. New Haven: Yale University Press.
- ↑ Prieto-Marques, A., Weishampel, D.B., Horner, J.R. (2006). "The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus". Acta Palaeontologica Polonica, 51 (1): 77–98.
- ↑ Holmes T (১৯৯৬)। Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science। Silver Burdett Press। আইএসবিএন 978-0-382-39147-7। ওসিএলসি 34472600।
- ↑ Salgado, L. & Gasparini, Z. (2006). "Reappraisal of an ankylosaurian dinosaur from the Upper Cretaceous of James Ross Island (Antarctica)". Geodiversitas 28 (1): 119–135.
- ↑ । ডিওআই:10.1126/science.264.5160.828।
|শিরোনাম=অনুপস্থিত বা খালি (সাহায্য) - ↑ Bakker, R.T. (1986). The Dinosaur Heresies. New York: William Morrow. আইএসবিএন ০-৮২১৭-৫৬০৮-৭ OCLC 363439291
- ↑ "Blood, tissue extracted from duck-billed dinosaur bone"। Cosmosmagazine.com। ২০০৯-০৫-০১। ২০১৩-১০-০৫ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০১৩-১০-০২।
- ↑ Wang, H., Yan, Z. and Jin, D. (১ মে ১৯৯৭)। "Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil"। Molecular Biology and Evolution। 14 (5): 589–591। ডিওআই:10.1093/oxfordjournals.molbev.a025796। পিএমআইডি 9159936। ২৮ আগস্ট ২০০৮ তারিখে মূল থেকে আর্কাইভ করা। সংগ্রহের তারিখ ২০০৭-১২-০৫।
- ↑ Chang BS, Jönsson K, Kazmi MA, Donoghue MJ, Sakmar TP (১ সেপ্টেম্বর ২০০২)। "Recreating a Functional Ancestral Archosaur Visual Pigment"। Molecular Biology and Evolution। 19 (9): 1483–1489। ডিওআই:10.1093/oxfordjournals.molbev.a004211। পিএমআইডি 12200476। সংগ্রহের তারিখ ২০০৭-১২-০৫।
- ↑ Schweitzer MH, Marshall M, Carron K, Bohle DS, Busse SC, Arnold EV, Barnard D, Horner JR, Starkey JR (১৯৯৭)। "Heme compounds in dinosaur trabecular bone"। Proc Natl Acad Sci U S A.। 94 (12): 6291–6। ডিওআই:10.1073/pnas.94.12.6291। পিএমআইডি 9177210। পিএমসি 21042 । বিবকোড:1997PNAS...94.6291S।
- ↑ Embery G, Milner AC, Waddington RJ, Hall RC, Langley MS, Milan AM (২০০৩)। "Identification of proteinaceous material in the bone of the dinosaur Iguanodon"। Connect Tissue Res। 44 (Suppl 1): 41–6। ডিওআই:10.1080/713713598। পিএমআইডি 12952172।
- ↑ Peterson, JE; Lenczewski, ME; Reed, PS (অক্টোবর ২০১০)। Stepanova, Anna, সম্পাদক। "Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs"। PLoS ONE। 5 (10): 13A। ডিওআই:10.1371/journal.pone.0013334। বিবকোড:2010PLoSO...513334P।
- ↑ "Dinosaur – Definition and More"। Merriam-Webster Dictionary। সংগ্রহের তারিখ ২০১১-০৫-০৬।
- ↑ Torrens, H.S. (১৯৯৩)। "The dinosaurs and dinomania over 150 years"। Modern Geology। 18 (2): 257–286।
- ↑ Breithaupt, Brent H. (১৯৯৭)। "First golden period in the USA"। Currie, Philip J. and Padian, Kevin (eds.)। The Encyclopedia of Dinosaurs। San Diego: Academic Press। পৃষ্ঠা 347–350। আইএসবিএন 978-0-12-226810-6।
- ↑ "London. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill." Dickens CJH (১৮৫২)। Bleak House। London: Bradbury & Evans। পৃষ্ঠা 1।
- ↑ Glut, D.F., and Brett-Surman, M.K. (১৯৯৭)। Farlow, James O. and Brett-Surman, Michael K. (eds.), সম্পাদক। The Complete Dinosaur। Indiana University Press। পৃষ্ঠা 675–697। আইএসবিএন 978-0-253-21313-6।
আরও পড়ুন
সম্পাদনা- রবার্ট টি. বাকের (১৯৮৬)। The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction। নিউ ইয়র্ক। আইএসবিএন 0-688-04287-2।
- হল্টজ্, থমাস অর. জুনিয়র (২০০৭)। Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages। নিউ ইয়র্ক: র্যান্ডম হাউস। আইএসবিএন 978-0-375-82419-7।
- পল, গ্রেগরি এস. (২০০০)। The Scientific American Book of Dinosaurs। সেন্ট মার্টিন'স্ প্রেস। আইএসবিএন 0-312-26226-4।
- পল, গ্রেগরি এস. (২০০২)। Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds। বাল্টিমোর: জনস হপকিন্স ইউনিভার্সিটি প্রেস। আইএসবিএন 0-8018-6763-0।
- স্টার্নবার্গ, সি. এম. (১৯৬৬). কানাডিয়ান ডাইনোসর, Geological Series, no. 54. Second ed. [Ottawa]: National Museum of Canada. 28 p., amply ill.
- Stewart, Tabori & Chang (১৯৯৭)। The Humongous Book of Dinosaurs। নিউ ইয়র্ক, স্টুয়ার্ট। আইএসবিএন 1-55670-596-4।
- Zhou, Z. (২০০৪)। "The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence" (পিডিএফ)। Naturwissenschaften। ৯১ (১০): ৪৫৫–৪৭১। ডিওআই:10.1007/s00114-004-0570-4। পিএমআইডি 15365634। বিবকোড:2004NW.....91..455Z। ২১ জুলাই ২০১১ তারিখে মূল (পিডিএফ) থেকে আর্কাইভ করা। সংগ্রহের তারিখ ৫ আগস্ট ২০১৪।
বহিঃসংযোগ
সম্পাদনা
- সাধারণ
- চিত্র
- The Science and Art of Gregory S. Paul Influential paleontologist's anatomy art and paintings
- Skeletal Drawing Professional restorations of numerous dinosaurs, and discussions of dinosaur anatomy.
- ভিডিও
- BBC Nature:Watch dinosaurs brought to life and get experts' interpretations with videos from BBC programmes including Walking with Dinosaurs ওয়েব্যাক মেশিনে আর্কাইভকৃত ১৯ আগস্ট ২০১৮ তারিখে
- BBC Explainer - Dinosaurs:Dinosaurs - a complete history in 4 minutes, animation
- জনপ্রিয়
- Dinosaurs & other extinct creatures: From the Natural History Museum, a well illustrated dinosaur directory.
- Dinosaurnews (www.dinosaurnews.org) The dinosaur-related headlines from around the world. Recent news on dinosaurs, including finds and discoveries, and many links.
- Dinosauria From UC Berkeley Museum of Paleontology Detailed information – scroll down for menu.
- LiveScience.com All about dinosaurs, with current featured articles.
- Zoom Dinosaurs (www.enchantedlearning.com) From Enchanted Learning. Kids' site, info pages and stats, theories, history.
- Dinosaur genus list contains data tables on nearly every published Mesozoic dinosaur genus as of January 2011.
- LiveScience.com Giant Dinosaurs Get Downsized by LiveScience, June 21, 2009
- প্রযুক্তিগত
- Palaeontologia Electronica From Coquina Press. Online technical journal.
- Dinobase A searchable dinosaur database, from the University of Bristol, with dinosaur lists, classification, pictures, and more.
- DinoData ওয়েব্যাক মেশিনে আর্কাইভকৃত ২০ ফেব্রুয়ারি ২০০৭ তারিখে (www.dinodata.org) Technical site, essays, classification, anatomy.
- Thescelosaurus! By Justin Tweet. Includes a cladogram and small essays on each relevant genera and species.